

Epoques Géologiques
CAMBRIEN (540-500 Ma)
L'environnement :
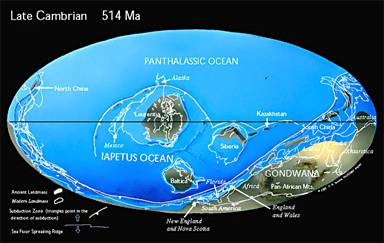
Le Cambrien commence alors que s'achève la dernière glaciation du protérozoïque, d'où une transgression générale, et une grande extension des mers épicontinentales, qui offrent ainsi des habitats nouveaux. Aucun continent n'est alors localisé sur un pôle, et le climat global aurait été relativement chaud. La glaciation suivante n'arrivera qu'à l'Ordovicien.
Quelques organismes caractéristiques :
La faune de Burgess Pass :
Le site de Burgess est une carrière de schistes, datés de 530 Ma. Les reconstructions paléo-environnementales qui en ont été faites indiquent que ces sédiments se sont déposés dans un bassin profond, sous la forme de coulées de boues glissant le long d'un escarpement de plusieurs centaines de m'ètres, depuis un récif algaire en bordure du continent.
Ce gisement a fourni une faune variée, dont certains représentants sont aujourd'hui rapprochés de phyla connus et encore actuels (Arthropodes, Annélides et Chordés), tandis que d'autres n'ont pu être rattaché à aucun phylum ultérieur, du moins pour l'instant.
Fossiles attribuables à des phyla connus :
|
Canadaspis, représentant des Arthropodes |
Marella, apparenté aux Trilobites |
||
|
|
Pikaia, premier représentant connu des Chordés |
||
|
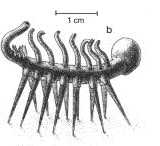
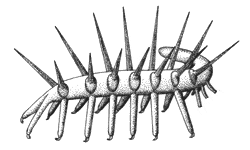
Le "cas" Hallucigenia : Ce fossile de Burgess, initialement reconstitué selon la représentation de gauche, ci-dessous, paraissait inclassable. Depuis la découverte, en Chine, d'autres organismes mieux conservés, d'age similaire, eux aussi vermiformes, au corps ornés de plaques osseuses portant des épines, et que l'on a reliés aux actuels Onychophores, une nouvelle reconstruction de Hallucigenia a été proposée, à droite : le fossile aurait initialement été reconstitué "à l'envers", et les épines prises pour des pattes. |
|||
|
|
|
||
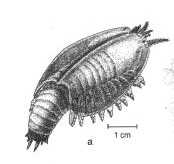

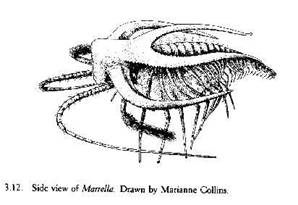

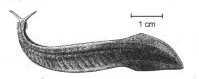
Fossiles d'affinité incertaine ou sans affinité avec les phyla actuels :
|
Anomalocaris, le plus grand prédateur de l'époque |
Opabinia, prédateur à cinq yeux et une pince préhensile centrale |
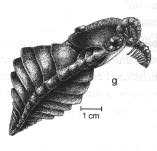

Les études phylogénétiques récentes placent ces fossiles, et d'autres, dans des clades spécifiques à la base du clade des Arthropodes.
Depuis la description de ces fossiles de Burgess, d'autres localités fossilifères datées du Cambrien ancien ont été découvertes et étudiées, en particulier en Chine. On y a décrit des organismes appartenant visiblement aux mêmes groupes de ceux de Burgess, occupant les mêmes habitats et les mêmes niches écologiques.
Dans l'ensemble, ces sites prouvent que la faune du début du Cambrien était déjà diversifiée et qu'elle occupait aussi bien l'intérieur du sédiments (vers...), l'interface eau-sédiment (petits Arthropodes...), la surface du sédiment [avec des formes mobiles (Arthropodes) ou fixées ("éponges"...)] que l'eau libre (Arthropodes, Chordés primitifs, Lobopodes...) . Certains de ces organismes étaient d'évidence carnivores. Ainsi, des écosystèmes relativement complexes, inconnus précédemment, étaient déjà en place il y a 530 Ma. (Cf. J. Vannier, Aux sources de la diversité du vivant, Pour la Science 323, Septembre 2004.)
Autres organismes des terrains cambriens
Les Radiolaires :
Ces Protistes Actinopodes, pourvus de symbiontes chlorophylliens (fréquemment des Dinoflagellés), se caractérisent par un test d'opale (silice amorphe), ponctuées d'orifices par lesquels émergent des pseudopodes. On en connaît deux classes, mais une seule, la classe des Polycystines, comprend des animaux au test suffisamment épais pour être fossilisable. Les premiers représentants de ce dernier groupe apparaissent au Cambrien ancien. Tous sont marins, et planctoniques.
Remarque annexe : Les Polycystines sont actuellement subdivisés en deux sous-ensembles, selon la forme de leur test : les Spumellaires, à symétrie radiale, et les Nacellaires, à symétrie bilatérale. Cependant, le classement des Radiolaires paléozo•ques dans l'un de ces groupes est quelquefois difficile.
Les Archaeocyathes :
|
Ce groupe, peut-être apparenté aux Démosponges, n'est connu qu'au Cambrien. Apparus il y a 530 Ma, ces organismes se diversifient très vite et constituent les premiers récifs de cette période.
Image : S. M. Stanley, Earth and Life through time, Freeman & Co.
|
|

Les Ostracodes :
Crustacés sans segmentation apparente, au corps entièrement contenu dans une carapace calcaire à deux valves. Ils apparaissent eux aussi dès le début du Cambrien. (Ci-contre, un Ostracode actuel) |
|

Apparition et diversification des Trilobitomorphes :
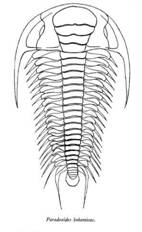
Les Trilobites Polymeridés Cambrien montrent bien les caractères plésiomorphes de ce groupe : la division en trois parties longitudinales (plèvre-rachis-plèvre) et transversales (Céphalon-Thorax-Pygidium), la glabelle, les lignes de suture.... Ils apparaissent au Cambrien et atteignent leur plus forte diversité à la fin de cette période (63 Familles ont été décrites au Cambrien récent, alors que ce nombre tombe à 19 dès le Silurien, pour se réduire à 4 au Carbonifère). Au cours de l'Ordovicien, ils présenteront des formes plus spécialisées, mais une diversité familiale en nette diminution.
|
Paradoxides |
Olenellus |
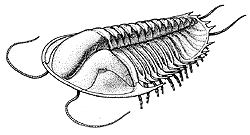
C'est aussi au Cambrien qu'apparaissent les Agnostidés, classe de Trilobites très probablement planctoniques, toujours de très petite taille et aveugles.
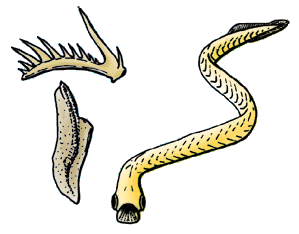
Apparition des Conodontes :
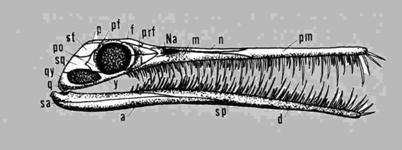
Les Conodontes n'ont longtemps été connus que sous la forme de petites pièces d'apatite dentelées, aux formes très variables. Ils apparaissent au Cambrien moyen, et ne disparaîtront qu'au Trias. Leur abondance et leur variété en ont fait d'excellents outils de biostratigraphie et de biochronologie pour les temps paléozoïques, bien que l'on ignorât à quel animal ils appartenaient. Ce n'est qu'à la suite de la description, dans des terrains du Carbonifère inférieur d'Edimbourg, d'empreintes d'un animal vermiforme comportant des conodontes, (un "animal-conodonte" d'une quarantaine de millimètres de long) et pourvu de myotomes, qu'on a pu rapprocher ces animaux des Chordés. Trois à quatre types de conodontes étaient associés dans l'animal pour former un appareil masticateur, qui devait lui permettre d'attraper, d'avaler et de déchiqueter ses proies.
Les "animaux-conodontes" sont maintenant regroupés dans la Classe des Euconodontes, et placés vers la base de l'arbre phylogénétique des Vertébrés, vu qu'ils synthétisaient, eux aussi, des structures en apatite.
Ils étaient traditionnellement associés à deux autres taxons, les Protoconodontes et les Paraconodontes, en un phylum des Conodontes, en dépit de leurs structures histologiques assez différentes. Pour l'heure, les spécimens pourvus d'appareils masticateurs articulés n'appartiennent qu'aux Euconodontes, et les deux autres groupes n'ont peut-être pas de lien direct avec ce dernier. (D'après P. Janvier, site Internet The Tree of Life, http://tolweb.org/tree/)
|
|
Reconstitution d'un Euconodonte, à droite, et schéma de deux conodontes, à gauche. Source : http://tolweb.org/tree/
|
ORDOVICIEN (500-435 Ma)
L'environnement :
La fin de l'Ordovicien est marquée par une extinction importante, qui touche ~ 25% de la faune marine. Elle pourrait être liée à l'arrivé du Gondwana au niveau du pôle Sud, qui s'accompagne de l'apparition d'une calotte glaciaire centrée sur l'actuel Sahara, et d'un changement dans la courantologie (développement d'une psychrosphère).
Quelques organismes caractéristiques :
Les graptolites :
Ces organismes coloniaux à "squelette" chitineux sont maintenant rattachés au phylum des Hémichordés et à la classe des Ptérobranches (grâce à l'étude détaillée de leur structure au microscope électronique). Ils sont apparus au Cambrien, avec des formes benthiques (Dendroïdes), pour s'éteindre au Carbonifère. Mais les formes planctoniques (Graptoloïdes) ne se rencontrent qu'à partir de l'Ordovicien. A l'inverse des Dendroides benthiques, formés d'un nombre importants de rameaux arborescents, plus ou moins reliés entre eux transversalement, les Graptoloides présentent des formes plus simples, avec un nombre de rameaux réduit, parfois un seul. Ils constituent une part importante et diversifiée du plancton marin durant l'Ordovicien et le Silurien. Pour ces deux époques, ils constituent ainsi d'excellents fossiles stratigraphiques.
|
Deux images du graptolite ordovicien Didymograptus, à droite en moulage tridimensionnel (la barre blanche représente 1 mm). |
|


Les brachiopodes :
Apparus au Cambrien, les brachiopodes atteignent dès l'Ordovicien un pic de diversité (en nombre de Genres) qu'ils ne retrouveront (et dépasseront) ensuite qu'au Dévonien.
Les trilobites :
L'Ordovicien ancien est encore une époque faste pour ce groupe, avec 61 Familles connues, chiffre qui descend à 42 à la fin de l'Ordovicien.
C'est à l'Ordovicien qu'apparaissent les trilobites capables de s'enrouler sur eux-mêmes (volvation), à la manière des cloportes ou des tatous actuels.
Epoque précédente / Retour en haut / Epoque suivante
SILURIEN (435-408 Ma)
L'environnement :
Le Silurien est une période de stabilisation du climat, marqué par la disparition des calottes glaciaires, qui se traduit par une transgression marine, et la mise en place, sur les plateaux continentaux ainsi immergés, de nombreux récifs.
Quelques organismes caractéristiques :
Organismes marins:
La radiation des Crinoïdes :
Les Crinoïdes, dont les premiers représentants pourraient remonter au Cambrien, présentent une radiation à partir du Silurien. Ils resteront dès lors largement représentés dans les faunes récifales jusqu'à la crise de la limite Permien-Trias. Après cet événement, ils se développeront de nouveau au Jurassique et au Crétacé, sans toutefois jamais revenir à la diversité qu'ils avaient atteinte au Paléozoïque.
Le succès des Brachiopodes :
Au Silurien puis au Dévonien, les Brachiopodes articulés présentent leur diversité la plus importante, en nombre de taxons d'ordre supérieur (Ordres) plutôt qu'en nombre de Genres ou d'Espèces, comme le montre la figure ci-dessous : tous les Ordres connus y sont représentés (Orthidés, Strophoménidés, Pentameridés, Rhynchonellidés, Spiriféridés, Térébratulidés).
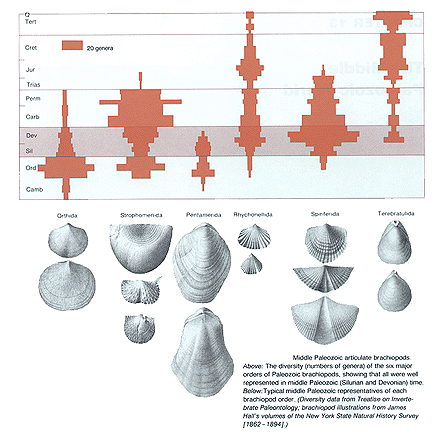
Image d'après S. M. Stanley, Earth and Life through time, Freeman & Co.
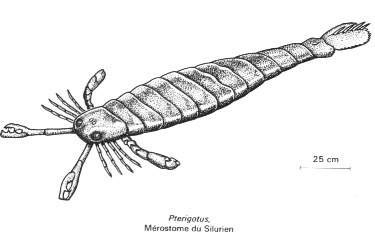
Les Euryptéridés :
Les Euryptéridés constituent un groupe éteint d'Arthropodes marins, apparus à l'Ordovicien et disparus au Permien, mais surtout abondants dans les terrains du Silurien supérieur et du Dévonien. Certains d'entre eux, les Ptérigotidés (ci-dessous, à droite Pterigotus), atteignaient un mètre de long, et représentaient probablement d'importants prédateurs des mers silurienne et dévonienne.
|
|
|
|
Eurypterus Image : UCMP Web Site |
Pterigotus Image : Encyclopaedia Universalis |

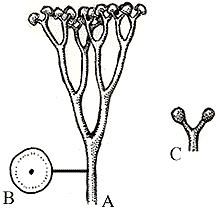
Les premiers végétaux aériens :
Des spores, éventuellement groupées en tétrades, ont été retrouvées dans des terrains datés de l'Ordovicien moyen au Silurien ancien (432 Ma). Cependant, les premières traces fossiles de végétaux vasculaires, à port dressé, datent de cette dernière période. Leurs formes sont simples, leur section cylindrique.
|
La plus ancienne forme végétale connue, nommée Cooksonia, a d'abord été découverte dans des terrains Dévonien du Pays de Galles. On en connaît maintenant des spécimens datant du Silurien terminal. Ils forment cependant un groupe paraphylétique.
A : Cooksonia caledonica, B : coupe montrant la protostèle. C : sporanges de Cooksonia hemisphaerica (Image : UCMP Web site)
Le plus ancien fossile attribué à une Lycophyte provient de terrains Silurien d'Australie (Baragwanathia longifolia, (~3 cm)).
Illustration ci-contre: Dossier Pour la science, La valse des espèces, Juillet 2000.
|
|

Epoque précédente / Retour en haut / Epoque suivante
DEVONIEN (408-355 Ma)
L'environnement :
Le Dévonien est marqué, sur les continents émergés, par l'arrivée des premiers tétrapodes semi-terrestres, mais aussi, pour les végétaux, à fin de cette période, par l'apparition des plantes à feuilles larges et aplaties, qui est fortement lié à l'évolution de l'atmosphère.
Note 1 :
Auparavant, tous les végétaux ne présentaient que des microphylles, c'est-à-dire des feuilles petites, coniques, semblables aux tiges. De plus, ces végétaux avaient un nombre de stomates par unité de surface relativement faible, ce que l'on pense corrélé à un taux élevé de CO2 atmosphérique (ces stomates relativement peu nombreux suffisaient dans ces conditions, à assurer les échanges gazeux, tout en empèchant une évapotranspiration excessive). Or, à partir du Dévonien, les plantes fossiles montrent une densité stomatique croissante : on en déduit qu'il s'est produit vers cette époque une chute du taux du taux de CO2 atmosphérique (de 300 à 30 Pa, en termes de pression partielle de CO2). Cette chute serait liée à une augmentation de l'érosion des minéraux silicatés et des carbonates, désormais altérés par les racines des plantes.
Mais des modélisations récentes des feuilles (Beerling et al., Nature 410, 15 Mars 2001) montrent que l'apparition des feuilles aplaties n'a pas été possible avant que les plantes ne disposent d'une densité stomatique assez importante : en effet, une densité stomatique faible, associée à une forme aplatie, sous des taux de CO2 encore élevés, auraient entra”né une élévation thermique en mème temps qu'une évapotranspiration accrue, aboutissant à des conditions létales pour la plante. Par conséquent, mème si, en termes de processus de développement et de capacité à synthétiser les tissus adéquats, l'apparition des feuilles larges paraissait possible bien avant la fin du Dévonien, les conditions climatiques et les besoins physiologiques des plantes n'auraient pas permis leur développement avant la fin du Dévonien.
Note 2 :
Le Dévonien est ponctué par plusieurs épisodes d'extinction, appelé événements, le plus important étant l'événement Kellwasser (du nom de la vallée allemande où se situe l'affleurement de référence qui montre ce niveau stratigraphique). Il marque la limite entre les deux derniers étages (et âges) du Dévonien, le Frasnien et le Famennien. Ces crises affectent surtout le benthos, en particulier les Foraminifères, mais aussi de nombreux autres groupes. Ces extinctions auraient des causes d'abord terrestres, peut-ètre accentuées par de petits impacts de corps extraterrestres : un réchauffement, entamé au Givétien, culminerait au Frasnien, et entra”nerait une dysoxie des eaux peu profondes des plateaux continentaux, immergés à la suite de transgressions marines successives. Au Famennien, au contraire, une importante régression s'ajoute à une chute des températures (peut-ètre en liaison avec la chute de la teneur atmosphérique en CO2).
Quelques organismes caractéristiques :
Organismes marins :
Coraux :
On peut retenir une forme caractéristique du Dévonien, appartenant aux coraux dits Rugueux (anciennement appelés Tétracoralliaires), Calceola sandalina (facilement reconnaissable par sa forme en "babouche")
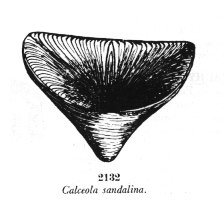
Image : A. Chavan & A. Cailleux, Détermination pratique des fossiles, Masson, 1980
Brachiopodes : l'apogée des Spiriféridés
Outre les Spiriféridés, le Dévonien est la période de plus forte diversité des Brachiopodes, avec un nombre de Genre maximal, et six Familles représentées.

image : UCMP Web Site
Arthropodes : Trilobites Polymeridés

Phacops est un trilobite caractéristique du Dévonien, aisément reconnaissable par sa glabelle énorme et hérissée de tubercules. On le trouve en abondance dans les terrains dévoniens du Maroc. (Figure : Phacops rana, site internet du Museum of Paleontology de Berkeley, www.ucmp.berkeley.edu)
Céphalopodes : Clyménidés
Cet ordre de Mollusque Céphalopode apparaît et se diversifie au Frasnien (Dévonien récent) pour s'éteindre au Famennien, lors des épisodes de crise de la limite Dévono-Carbonifère. A la différence des Ammonitidés, auxquels ils ressemblent, Les Clyménidés ont une coquille légèrement allongée, et non circulaire, et leur siphon est situé sur la face dorsale de la coquille, et non en position ventrale (cas des Ammonitidés) ou centrale (cas des Nautiloïdés).
Leur succès puis leur déclin très rapide en font d'excellents fossiles stratigraphiques, avec une résolution temporelle pouvant atteindre 1 Ma seulement.
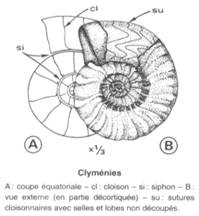
Figure : Foucault & Raoult, Dictionnaire de géologie, Masson
Les poissons cuirassés :
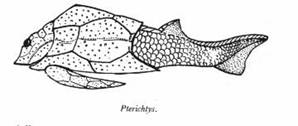
On connaît au Dévonien un grand nombre de "poissons", avec ou sans mâchoire (comme Pterichtys, ci-dessous à gauche), portant une armure de plaques dermiques au niveau du crâne, du thorax et pour certains sur les nageoires pectorales.
Les poissons cuirassés, et à mâchoires, groupe-frère probable des Gnathostomes, sont appelés Placodermes (Placodermi). [Le plus ancien connu date du Silurien]. Certains atteignaient de très grandes tailles, en particulier Dunkleosteus, ci-dessous. Aucun n'avait de dents véritables, uniquement des plaques dermiques pointues et aiguisées. Par contre, les vrais Chondrychtiens, entre autres les vrais Requins, apparaissent à la mème époque. Les Placodermes s'éteignent à la fin du Dévonien.
|
Pterichtys, poisson cuirassé sans mâchoire Figure : A. Chavan & A. Cailleux, Détermination pratique des fossiles, Masson
|
Figure: Stanley, S. M., Earth and Life through Time, Freeman. |

Organismes terrestres et "sortie des eaux"
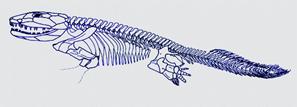
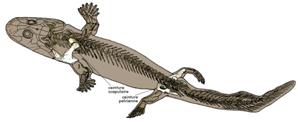
Premiers tétrapodes :
C'est du Dévonien que sont datés les fossiles les plus célèbres de Tétrapodes primitifs, dont les deux plus fameux, Ichtyostega et Acanthostega.
|
|
|
|
Ichtyostega, probablement capable de ramper sur les berges avec ses pattes avant. |
Acanthostega, incapable de se mouvoir hors de l'eau, vu l'orientation de ses membres, qui lui permettait peut-ètre de se stabiliser dans l'eau des ruisseaux pour chasser à l'affût. |
CARBONIFERE (355-295 Ma)
L'environnement :
|
|
|
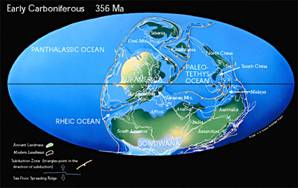
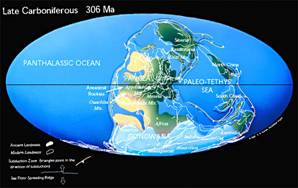
Les sédiments des continents alors situés dans l'hémisphère sud permettent de montrer que celui-ci était englacé (d'où des alternances de transgressions et régressions avec les fluctuations de taille de ces calottes), ce qui pourrait être lié à la baisse de la teneur atmosphérique en CO2, entamée au Dévonien (cf. fiche Dévonien). Dans l'hémisphère Nord, le climat était probablement chaud et humide, avec des forêts tropicales et subtropicales au niveau de l'actuelle Amérique du Nord, de l'Asie et de l'Europe. Cette végétation sera à l'origine des vastes gisements de charbons qui caractérisent aujourd'hui les séries sédimentaires de cette époque. La rareté des cernes de croissances dans ces restes végétaux suggère un climat sans saisons marquées.
Quelques organismes caractéristiques :
Dans les mers, les fonds sont peuplés par de nombreux genres de brachiopodes (voir la figure de la fiche Silurien : en nombre de Genres, ce groupe est à son apogée), ainsi que par des Crinoïdes; les trilobites se raréfient (il n'en reste que quatre Familles), alors que les Foraminifères abondent. Les Poissons cuirassés disparaissent avec les épisodes de crise de la fin du Dévonien, au profit des Chondrichtyens (poissons cartilagineux) et des Ostéichtyens (poissons osseux).
Végétaux :
|
Ils sont nombreux. Ce sont surtout de Ptéridophytes, souvent arborescentes (Lycopodiacées, Sphénophytes (Equisétinées), Filicinées), et de "Fougères à graines" (Ptéridospermales), comme Neuropteris. (échelle 1/2)
Image : S. M. Stanley, Earth and life through time, Freeman |
|
|
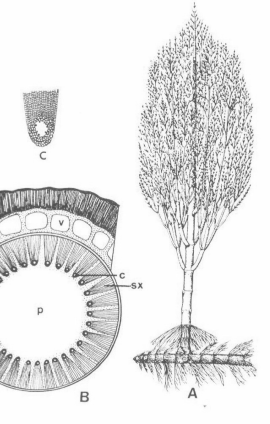
Parmi les Sphénophytes, on peut citer, au Carbonifère récent, des fossiles comme Calamites, ci-contre, un arbre d'une trentaine de mètres, dont on trouve souvent des moules internes dans les couches de houille. Rq : Ses différentes parties (feuilles, racines, et tronc) avaient été décrites sous des noms différents, avant qu'on les interprète comme issues d'une même plante.
Toutes les images de paléobotanique proviennent du site Virtual Paleobotanical Lab de l'Université de Berkeley. |
 |
|
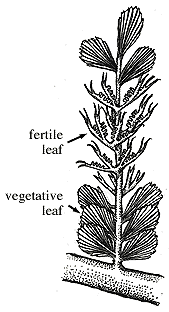
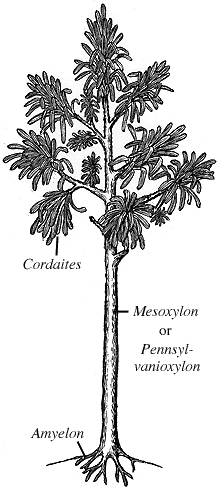
Les Progymnospermes, apparues au Dévonien, sont aussi connues au Carbonifère inférieur, avec par exemple, Archaeopteris ("feuille ancienne"). Là encore, le nom désignait initialement la seule feuille, avant que l'on attribue un tronc fossile, Callixylon ("bel arbre") à la même plante. |
 |
|
Les Progymnospermes, apparues au Dévonien, sont aussi connues au Carbonifère inférieur, avec par exemple, Archaeopteris ("feuille ancienne"). Là encore, le nom désignait initialement la seule feuille, avant que l'on attribue un tronc fossile, Callixylon ("bel arbre") à la même plante |
 |
Céphalopodes :
|
C'est au Carbonifère que l'on trouve le genre Goniatites, ci-dessous, représentant caractéristique de l'Ordre des Goniatidés, apparus au Dévonien et qui s'éteindront à l'occasion de la crise Permo-triasique. Image : A.Foucault & A.Raoult, Dictionnaire de Géologie, Masson |
|

Arthropodes :
|
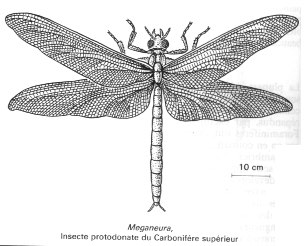
Ils abondent dans les forêts carbonifères, avec parfois de grandes tailles par rapport aux arthropodes aériens actuels. Les insectes ailés seraient apparus à la fin du Dévonien. Parmi les Insectes, l'une des plus célèbres est sans doute la "libellule géante" Meganeura, d'une envergure de 70 cm (l'équivalent d'un Goeland).
Image : Encyclopaedia Universalis |
|
Tétrapodes :
|
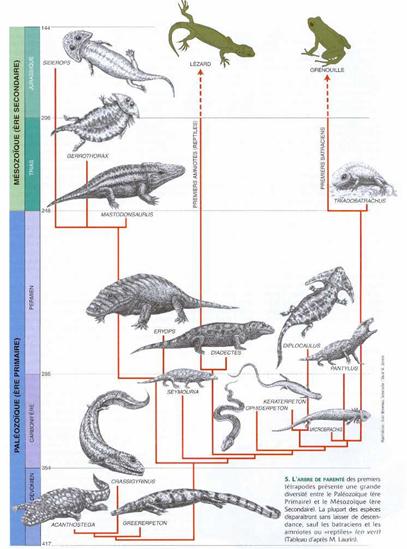
Les Amphibiens étaient présents au cours du Carbonifère, à la fois sur la terre ferme et dans les eaux douces. Certains, comme Crassigyrinus, (cf. figure), étant "revenus à l'eau" avec une nouvelle atrophie de leurs membres. Ci-contre, un arbre phylogénétique des premiers tétrapodes. Notez la radiation de ces animaux au début du Carbonifère. Par contre, c'est aussi au Carbonifère, peut-être dès le Carbonifère ancien, qu'apparaissent les représentants du "grade évolutif Reptile", c'est-à-dire les animaux à oeuf amniotique (d'après la découverte en 1988 d'un fossile de cette date, attribué à un Reptile, dans des terrains d'Ecosse). Malheureusement, on n'a pour l'heure quasiment pas de fossiles datant du carbonifère ancien, et donc à même de documenter le passage des formes dévoniennes (Ichtyostega et Acanthostega) aux amphibiens du Carbonifère récent. |
Source: Dossier Pour la Science, La valse des espèces, Juillet 2000 |
Epoque précédente / Retour en haut / Epoque suivante
PERMIEN (295-250 Ma)
L'environnement :
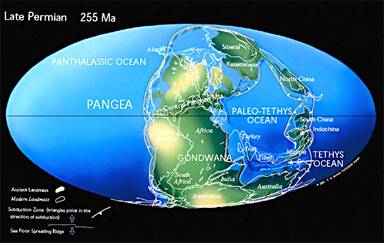
Les continents permiens sont rassemblés sur une face de la planète, quasiment d'un pôle à l'autre, en une Pangée, entourée d'une Panthalassa. On peut cependant encore individualiser un bloc Gondwana au sud, et la Laurasia au nord.
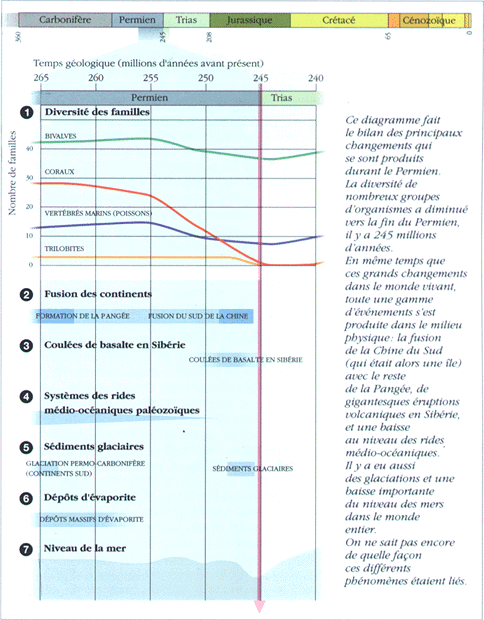
La fin du Permien correspond à la plus importante des extinctions en masse connue. Elle aurait affecté, selon certains auteurs, jusqu'à 96 % des espèces marines ! Elle paraît responsable, directement ou en tant que "coup de grâce", de l'extinction totale de certains grands groupes, aux modes de vie parfois très différents. Parmi eux les Coraux Tabulés et Rugueux, des Foraminifères (les Fusulinidés), les Trilobites, des Echinodermes, les Blastoïdes, des Mollusques, les Goniatitidés.
Nombres d'autres groupes voient leurs effectifs et leur diversité considérablement affectés, soit au cours du Permien, soit à la limite même : ainsi 75% des Familles de Bryozoaires disparaissent, 50% des Familles de Brachiopodes, soit 90% des Genres, s'éteignent au cours du Permien récent, un seul Genre d'Echinides (Oursins), Miocidaris, survit et est donc à l'origine de l'ensemble des Echinides qui suivront. Sur la terre ferme, les "reptiles mammaliens" sont également très fortement touchés.
Enfin, la limite Permien/Trias se caractérise dans les sédiments par un "événement fongique" : les faciès de mers peu profond montrent, juste au dessus de la limite P/T, un fin niveau très riches en restes de champignons et en débris plus ou moins carbonisés de plantes terrestre, surmonté de niveau riches en pollens de Lycopodes. Cet ensemble suggère un épisode de destruction importante de la flore, dont les restes auraient été utilisés par les champignons saprophytes, avant la re-colonisation végétale, assurée d'abord par les Lycopodes.
Ces extinctions, dans la mesure où elles ne sont pas toutes brutales, ont très probablement plusieurs causes interconnectées.
- La formation de la Pangée a réduit la surface des mers épicontinentales, très peuplées, et s'est traduit par une régression marine sans précédent, et, sur le continent, par un climat très sec et à forte saisonnalité (importants dépôts d'évaporites dans les formations du permien supérieur d'Amérique du Nord et d' Europe).
- Ces masses de sels séquestrées sur les continents aurait entraîné, qui plus est, une baisse de la salinité de l'eau de mer, affectant les organismes marins sténohalins.
- La disparition des calottes glaciaires présentes, au Permien Ancien, sur la partie Sud de la Pangée.
- Les trapps de Sibérie, datés de la limite Permien-Trias, signent un épisode de volcanisme d'une intensité inconnue depuis, dont les conséquences climatiques ont sérement été importantes.
En 2002 ont été publiées des datations de basaltes enfouis à 4 km de profondeur, atteints au moyen de carottages, à 1000 km à l'Ouest de la zone d'extension précédemment admise pour les trapps : Ces laves appartiendraient aussi aux trapps sibériens, ce qui signifie que la surface couverte par ces laves atteindrait 3,9 millions de km2 au lieu des 2 millions estimés auparavant (Reichow et al.(2002), Science, v.296, 1846-1849).
- Enfin, l'hypothèse d'un impact météoritique, un temps écartée au profit d'une explication climatique et volcanique, a récemment été confortee par de nouvelles etudes:
Des fullerènes (molécules à 60 atomes de carbone en forme de "ballons" creux), extraits de couches datées de la limite P/T dans des affleurements de Hongrie, de Chine et du Japon, contiennent des concentrations inhabituelles de molécules d'Argon et d'Hélium, emprisonnées lors de la formation des fullerènes, et avec des valeurs isotopiques similaires à celles des chondrites carbonées (Becker et al. (2001), Science v. 291, 1530-1533).
D'autres travaux, sur des terrains de Chine, ont montré l'existence d'un pic de Strontium et de Soufre dans les couches datant de cette limite (Kaiho et al. (2001), Geology v.29, 815-818), avec là encore des signatures isotopiques compatibles avec celles de matériaux extraterrestres.
|
Récapitulatif des données de la fin du Permien : |
|
|
|
Source: S. J. Gould (dir.), Le livre de la Vie, 1993, Seuil |
Quelques organismes caractéristiques :
Organismes marins :
Foraminifères : Les Fusulinidés
Apparus au Carbonifère, ces Foraminifères à test, dit "pseudofibreux", fréquemment en forme de fuseau, pourtant florissants au Permien, sont totalement anéantis par la crise Permien-Trias.
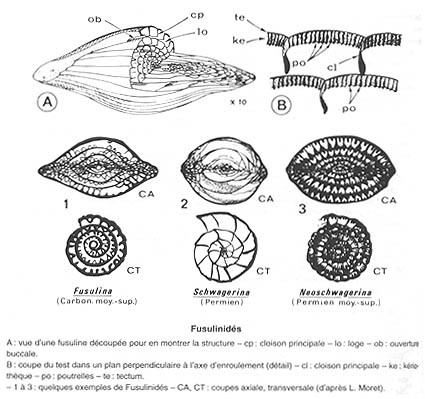
Source : A. Foucault & J-F. Raoult, Dictionnaire de géologie, Masson
Organismes terrestres :
Amphibiens :
Le Permien a livré un certain nombre de fossile d'Amphibiens (voir la figure de la fiche Carbonifère). On mentionnera juste ici, pour sa forme caractéristique, Diplocaulus, du Permien Ancien d'Amérique du Nord.
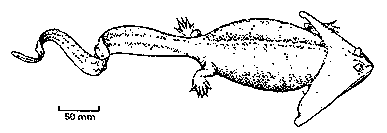
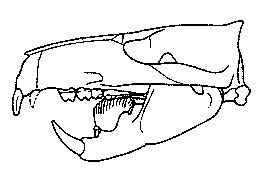
Les "reptiles" Synapsides :
|
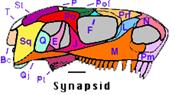
Pour mémoire, on désigne par ce nom les amniotes dont le crâne (schéma ci-contre) ne présente qu'une seule fenêtre temporale, entourée par les os appelés postorbital (po), jugal (j) et squamosal (sq). Ce groupe monophylétique inclut les "reptiles mammaliens" et les Mammifères. |
|
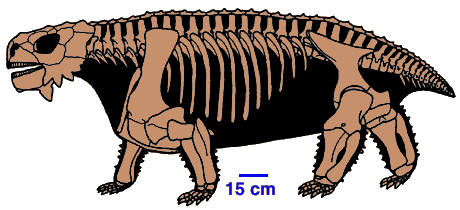
- Avec six familles représentant plus de 70% de tous les genres de Synapsides du Permien ancien, les Pélycosaures (Ordre des Pelycosauria (Benton, 2000)) constituait le groupe de tétrapodes le plus diversifié de cette période. Trois familles au moins remontent au Carbonifère Récent, mais leur diversité maximale n'est atteinte qu'au Permien.
|
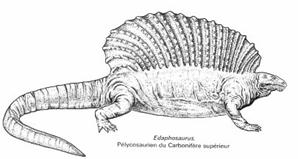
Dans deux familles de Pélycosaures, les Edaphosauridés (herbivores, comme Edaphosaurus, ci-dessous à gauche) et les Sphénacodontidés (carnivores, comme Dimetrodon, ci-dessous à droite), sont apparus des formes qui présentaient un fort allongement des épines dorsales, très probablement reliées entre elles par une membrane de peau en une "voile dorsale". L'étude de cette structure a montré que sa taille était plus fonction de la masse de l'animal que de sa seule longueur, et qu'elle était bien vascularisée, ce qui permet de penser qu'elle avait un rôle physiologique, probablement de thermorégulation : des calculs ont ainsi établi qu'elle permettait à un Dimetrodon de 250 kg de passer d'une température de 25 à 30°C en trois heures d'exposition au soleil au lieu de 12h en l'absence de cette structure. |
|
|
|
|
|
Edaphosaurus, ~2,5 m de long Image : Encyclopeadia Universalis |
Dimetrodon, ~3 m de long |

Rq : Cette morphologie est manifestement apparue trois fois indépendamment à cette époque : chez ces deux groupes de Pélycosaures, mais aussi chez un Temnospondyle (groupe de tétrapodes "Amphibiens" primitifs, apparus au Carbonifère) contemporain.
- Au Permien Récent, on trouve un autre groupe de Synapsides, l'Ordre des Thérapsides, dérivés des Pélycosaures. Ce groupe comprend des représentants fossiles qui présentent un certain nombre de traits "mammaliens", et les Mammifères. Les formes fossiles sont classées dans plusieurs sous-Ordres, dont les principaux sont indiqués sur la figure ci-dessous. Les plus connus ont été découvert dans les gisements Permo-Triasique du Karoo, en Afrique du Sud.
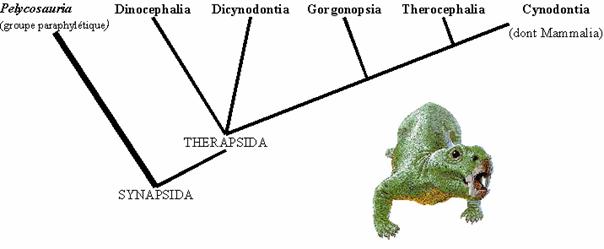
Cladogramme d'après M.J. Benton, Vertebrate Paleontology, Blackwell Science
A côté du cladogramme, une reconstitution d'un représentant des Dicynodontes, Diictodon, du bassin du Karoo.
Le groupe des Thérapsides comprenait des animaux de taille et de régime alimentaire très variable, depuis les petits carnivores coureurs de 30 cm jusqu'aux énormes herbivores de 5 m de long.
"Reptiles" Anapsides et Diapsides :
Ces deux autres catégories complètent le clade des Amniotes. Comme pour les Synapsides, leur définition repose sur la morphologie crânienne :
|
Les Anapsides sont dépourvus de fenêtre temporale (état plésiomorphe), et sont aujourd'hui représentés par les Tortues terrestres et marines... |
|
|
...alors que les Diapsides, qui comprennent aujourd'hui les lézards, les serpents, les crocodiliens et les oiseaux, en possèdent deux. Rq : les couleurs des os suivent le même code que l'illustration précédente de l'état Diapside. |
|

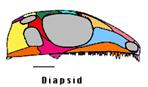
Source des figures : The Tree of Life Web Project.
- Les Anapsides remontent au Carbonifère, mais sont surtout représentés au Permien et au Trias, avec plusieurs groupes, dont. la position systématique par rapport aux Tortues actuelles (Testudina) est encore débattue.
Ci-dessous, un Scutosaure, membre des Pareiasauria, l'un des deux groupes proposés comme groupe-frère des Testudina. Certains chercheurs suggèrent que les plaques dermiques de ces animaux massifs pourraient avoir abouti à la carapace des Tortues.
Source: UCMP Web site
Un deuxième groupe actuellement classé parmi les Anapsides, en groupe-frère des testudines, les "Parareptiles" comprend au Permien des animaux revenus à un mode de vie entièrement aquatique, comme Mesosaurus, ci dessous.
|
|
Retrouvé en Afrique et au Brésil, l'animal, qui nageait probablement en eau douce ou dans des mers peu profondes, mesurait environ un mètre ou moins. Il présentait une longue queue aplatie en nageoire, qui assurait sa propulsion, et des pattes arrière aux doigts très longs, probablement palmés.
Le museau allongé, garni de nombreuses dents fines et pointues qui s'entrecroisent, constitue une adaptation à un régime piscivore, à l'instar des gavials actuels..
|

- Les Diapsides, dont les premiers représentants remontent au Carbonifère Récent, restent peu diversifiés au Permien Ancien, et présentent une radiation évolutive au Permien Récent.
|
Le plus remarquable de ces Diapsides du Permien Récent de Madagascar, est Coelurosauravus, premier vertébré "planeur" : ses vertèbres, allongées dans le plan horizontal, formait vraisemblablement une membrane qui lui permettait d'effectuer des vols planés, et sans doute de chasser ainsi les insectes volants. |
|
|
|
|
|
Image : M. J. Benton, Vertebrate Paleontology, 2000, Blackwell Science |
Image : S. J. Gould (dir.) Le livre de la vie, 1994, Seuil
|

Végétaux terrestres :
La glaciation du début du Permien et les conditions climatiques qui suivent favorisent l'expansion des plantes à graines (Ginkgoales et Gymnospermes) au détriment des grandes Lycopodes, Fougères et Prêles du Carbonifère.
Epoque précédente / Retour en haut / Epoque suivante
TRIAS (250-203 Ma)
L'environnement :
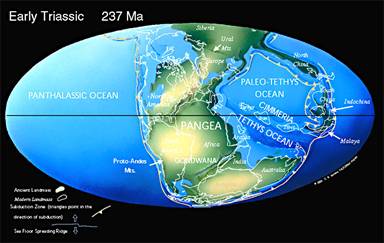
En termes d'environnement, le Trias ne diffère pas beaucoup du Permien. Les continents sont encore regroupés. Le climat général est chaud, avec un faible gradient Equateur-Pôles. Il n'y a pas de calotte polaire.
Par contre, suite à la crise qui constitue la limite Permien-Trias, les faunes sont fortement remaniées. On y constate la persistance de représentants des quelques groupes Permien survivants, et le développement de groupes nouveaux, qui se répandent et se diversifient en occupant les niches écologiques devenues vacantes.
Quelques organismes caractéristiques :
Organismes marins :
Coraux : Les Scléractiniaires
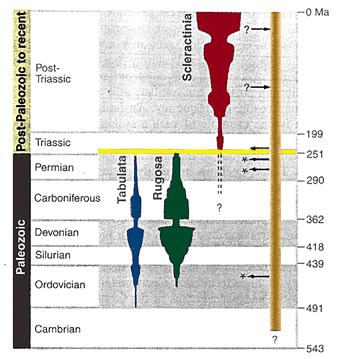
Les coraux calcifiés sont l'une des principales victimes de l'extinction Permien-Trias : Les deux groupes dominants du Paléozo•que, les Tabulés et les Rugueux, s'éteignent totalement à ce niveau. Les 14 premiers millions d'années du Trias ne fournissent aucun fossile de corail calcifié. Ce n'est que passé ce laps de temps que l'on trouve des représentants d'un nouveau groupe de corail, les Scléractiniaires, dont font partie les coraux actuels. Cette apparition brutale, et la difficulté à rapprocher phylogénétiquement ces animaux des groupes calcifiés précédents, fait penser que les Scléractiniaires auraient eu un ancêtre non calcifié, et par conséquent absent du registre fossile.
L'acquisition, ou la perte de squelette calcifié, se serait produite plusieurs fois au cours des temps géologiques, en relation avec la teneur en carbonate de calcium de l'eau de mer. Les conditions du début du Trias, auraient conduit à un changement de la chimie de l'océan, diminuant la précipitation de CaCO3. L'émergence des Scléractiniaires dans le registre fossile correspondrait au retour des conditions favorable à cette précipitation.
|
|
Coraux et apparentés au cours du temps : La longueur verticale des figures colorées montre l'extension temporelle des trois Ordres de coraux calcifiés, et leur épaisseur représente leur abondance et leur diversité relative. La ligne jaune à la base du Trias indique la période d'absence de coraux. La barre orange verticale sur la droite représente les formes sans squelette, pour lesquelles les fossiles sont rares ou inexistants. La figure n'implique donc aucune continuité des lignées au sein des groupes, ni que les formes sans squelette soient étroitement apparentées. Les astérisques indiquent l'apparition de sclératiniomorphes calcifiés. Les flèches vers la droite précisent les épisodes de perte de squelette calcifié, celle vers la gauche les épisodes de gain.
Figure et légende adaptées de Stanley & Fautin, The origin of modern corals, Science 291, 9 Mars 2001 |
Céphalopodes: Les Cératitidés
Ces Céphalopodes, typiques du Trias, succèdent aux Goniatitidés Permo-Carbonifère. Ils présentent des sutures assez simples, mais lobées et non anguleuses comme celles des Goniatites, avec un une "selle" orientée vers l'ouverture de la coquille, et un "lobe" légèrement dentelé orienté vers l'arrière. Ci-contre, un exemple de ces fossiles, sur lequel on a souligné la ligne de suture. |
|

Organismes terrestres :
Amphibiens :
Le Trias de Madagascar a fourni des fossiles des premiers représentants des groupes d'Amphibiens actuels, en particuliers les premiers Anoures, avec la grenouille Triadobatrachus (littéralement "grenouille du Trias")
Synapsides Dicynodontes :
Les "reptiles mammaliens" sont les principales victimes de la crise Permo-Triasique pour les faunes de vertébrés terrestres. Au début du Trias, on ne rencontre quasiment plus qu'un seul Genre de Dicynodonte, de taille moyenne, Lystrosaurus (ci-dessous).
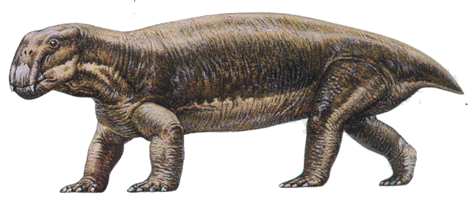
Source : Science & Vie, hors-série Dans le secret des mondes disparus, Décembre 2000.
Remarque : cet animal est fréquemment cité comme argument paléontologique en faveur de la tectonique des plaques et de l'existence du Gondwana, dans la mesure où l'on en a retrouvé des représentants sur plusieurs continents.
Cependant, ce groupe se diversifie à nouveau rapidement au cours du Trias, et l'on retrouve, 5 Ma après la crise, de grands Dicynodontes herbivores.
Synapsides Cynodontes :
(se reporter au cladogramme de la fiche Permien)
Alors que les Dicynodontes se diversifient à nouveau, le groupe des Cynodontes ("dents de chiens"), présente une radiation évolutive. Les représentants triassiques de ce groupe, qui comprendra les Mammifères, présentent déjà plusieurs caractères mammaliens, en particulier :
- Un palais secondaire rigide, qui sépare la bouche des conduits nasaux.
- Une mâchoire inférieure constitué d'un seul os, le dentaire.
- Une arcade zygomatique large, à travers laquelle passe des muscles masticateurs puissants.
- Des traces de nerfs et de vaisseaux au niveau du museau permettent de supposer la présence, chez ces Cynodontes, de vibrisses, et donc de poils, ce qui laisse aussi penser que ces animaux étaient endothermes.
Voici par exemple, ci-dessous, une reconstitution de Thrinaxodon, du début du Trias. Observez la mâchoire, constituée du seul os dentaire, et les dents postérieures, qui ne sont plus simplement coniques, mais portent trois cuspides. l'animal est représenté couvert de pelage, même si on n'en a pas de preuves directes.
|
|
|
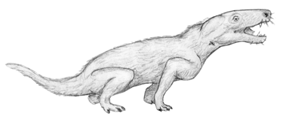
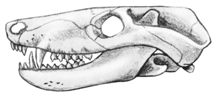
Images :Pr. Paul E. Olsen, http://www.ldeo.columbia.edu/dees/courses/v1001/dinos.2002.html
|
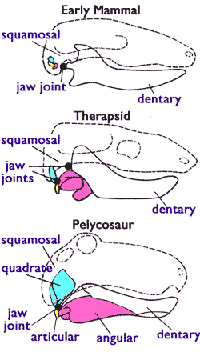
Les Cynodontes les plus récents permettent aussi de documenter le passage de l'articulation reptilienne de la mâchoire, à l'articulation mammalienne :
Un certain nombre de Cynodontes (indiqué ci-contre par "Thérapsides") présentaient les deux articulations simultanément, toutes deux fonctionnelles. Les formes fossiles de la fin du Trias deviennent ainsi difficiles à classer, entre Cynodontes "de grade reptile" et Mammifère vrais. |
 |
Anapsides :
C'est à la fin du Trias qu'apparaissent les premiers représentants des Testudinés, les vraies Tortues.
Ils ne présentent pas encore l'articulation des vertèbres cervicales qui permettra à leurs descendants de replier leur cou à l'intérieur de la carapace. Ce caractère n'apparaît qu'à partir du Jurassique, selon deux modalités qui distinguent les deux sous-ordres de Testudines :
- les Pleurodires, sans doute le type le plus ancien, dont le cou se plie dans le plan horizontal
- Les Cryptodires, chez qui le repliement s'effectue dans le plan vertical.
Diapsides :
Dans ce clade se développent deux sous-clades :
- celui des Lépidosauromorphes, qui comprendra lui-même deux ensembles de reptiles marins :
- les Ichtyosaures
- l'ensemble (Placodontes + Nothosaures + Plesiosaures), ou Sauropterygiens.
- celui des Archosauromorphes, qui regroupe :
- des formes essentiellement triassiques (Trilophosauridés et Rhynchosauridés)
- les Prolacertiformes
- et le groupe des Archosauria, dans lequel on rassemble les Ptérosaures, les Dinosaures (et les Oiseaux), et les Crocodiles.
Ainsi, parmi ces reptiles, beaucoup, connus dès le début du Trias, sont redevenus aquatiques. Ce retour a sans doute été entamé au Permien , car les conditions du Trias ne se distinguent pas suffisamment de celles de la période précédente, du point de vue de la disponibilité de nourriture marine (Brachiopodes, Céphalopodes, Bivalves, Poissons, etc·) pour que l'on puisse considérer cette brusque abondance de reptiles marins comme une conséquence de la crise Permien-Trias.
On présente ci-dessous quelques formes triasiques remarquables de ces animaux :
- Les Ichtyosaures, connus du Trias au Crétacé, présentent l'adaptation la plus poussée à la vie marine en haute mer, et sont un exemple remarquable de convergence évolutive avec les Poissons puis les Cétacés (cette convergence s'explique aisément par l'identité des contraintes hydrodynamiques du milieu, qui ont chaque fois favorisé les mêmes réponses adaptatives : corps fuselé, nageoire caudale propulsive, nageoire dorsale stabilisatrice, nageoires latérales assurant les changements d'orientation).

Image: Pour la Science
- Les Sauroptérygiens présentent un tous corps allongé, propulsé essentiellement par les mouvements des membres.
- Les Placodontes du trias moyen ont un corps massif, des dents très plates (d'où leur nom) et des incisives en spatule, probable adaptation à la consommation des mollusques accrochés aux rochers. Certains avaient un corps très large et aplati, semblable à celui d'une tortue, ce qui a amené certains chercheurs à rapprocher ce groupe des Testudines. Ci-dessous, une reconstitution de Placodus gigas.

- Les Nothosaures, connus au Trias moyen en Europe (Mushelkalk), montrent un corps et un cou allongé, avec une tête proportionnellement petite. Leur taille allait de 20 cm à plus de 4 m. Nothosaurus, ci-dessous, en est le représentant le plus typique.

- Les Plésiosaures, qui n'apparaissent qu'à la fin du trias, ressemblent beaucoup aux Nothosaures, dont ils descendent probablement. Généralement plus grand que ces derniers, leur adaptation à la nage est aussi encore plus poussée, avec des membres latéraux transformés en palettes natatoires attachés à des ceintures renforcées. C'est sous la forme d'un plésiosaure qu'est classiquement représenté l'hypothétique "monstre du Loch Ness".
- Chez les Prolacertiformes, on trouve au Trias un animal remarquable par son cou très allongée en comparaison du reste du corps, Tanystropheus. On lui attribue un mode de vie proche de l'eau, et un régime piscivore, ce cou démesuré lui permettant de pêcher depuis la berge.
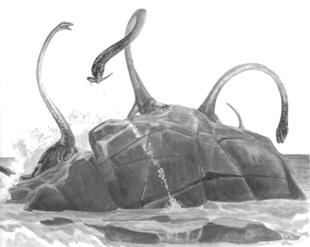
Image : S.J. Gould (dir.), Le livre de la Vie, Seuil
- Les premiers Dinosaures font leur apparition au trias moyen. Ils présente une innovation majeure, la posture érigée (les membres sont tendus à la verticale sous le corps et non plus arqués sur les côtés) qui évolue chez certaines formes en une posture bipède. Leur taille et leur diversité restent néanmoins modestes durant tout le Trias.
Les bouleversements de la fin du Trias :
Les faunes et les flores de la fin du Trias présentent des changements profonds, suffisant pour que l'on puisse parler de crise fini-triasique.
(source des chiffres donnés ci-après : F. Lethiers, Evolution de la biosphère et événements géologiques, Gordon and Breach Science Publishers).
- Dans les mers, 76% des espèces disparaissent, soit 23 % des familles. Parmi les grands groupes affectés figurent :
- Les Conodontes, qui disparaissent entièrement.
- Les Orthocéridés, céphalopodes à coquille externe droite (ortho-) et, parmi les Ammonoïdés, tous les Cératitidés, s'éteignent également durant cet épisode.
- Les récifs coralliens régressent, plusieurs formes constructrices (éponges et scléractiniaires) s'éteignent.
- Les Bivalves sont particulièrement touchés en nombre d'espèces éteintes, mais aucune Famille ne disparaît.
- De nombreuses espèces de Reptiles marins sont également touchées.
- Sur les continents, 22% des familles de Tétrapodes s'éteignent. On y enregistre surtout un changement faunique important à la limite Carnien-Norien (Trias récent).
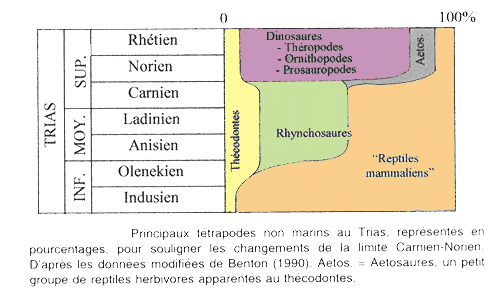
Figure : d'après F. Lethiers, Evolution de la biosphère et événements géologiques, Gordon and Breach Science Publishers
Les causes de la crise sont toujours en débat.
Un travail récent (Science 296, 17 May 2002) portant sur un grand nombre de sites fossilifères Nord-américains, a étudié les modifications des traces fossiles de Tétrapodes autour de la limite Trias-Jurassique. Il montre une chute du nombre, et de la diversité, des traces attribuées aux non-dinosauriens après cette limite, et à l'inverse, une augmentation de taille et de diversité des traces assignées aux Dinosauriens.
Les auteurs de cette étude trouvent également sur certains sites un pic d'abondance de spores et de pollen, et une anomalie positive dans la teneur en Iridium des sédiments, à la limite Trias-Jurassique.
Ces résultats leur permettent donc d'avancer :
- d'une part, qu'un impact météoritique serait l'une (au moins) des causes de cette crise,
- et, d'autre part, que ce bouleversement a favorisé l'expansion et la domination ultérieure du groupe des Dinosauria, autrement dit que l'expansion des Dinosaures au détriment des autres groupes de Tétrapodes ne résulte pas seulement d'un remplacement par compétition mais (au moins en partie) de l'occupation d'un espace écologique préalablement "nettoyé" par un épisode d'extinction.
JURASSIQUE (203-135 Ma)
L'environnement :
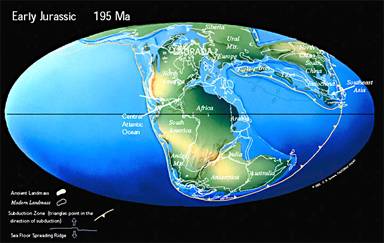
Sec et aride au centre de la Pangée au début du Jurassique, avec des zones tropicales et subtropicales plus étendues qu'aujourd'hui, un climat tempéré au hautes latitudes, et l'absence de calotte polaire, le climat se modifie avec la fragmentation progressive du supercontinent. L'Atlantique Nord commence à s'ouvrir, alors que la Téthys poursuit son expansion vers l'Ouest. Le Jurassique récent présente des climats moins secs sur l'intérieur des terres, tandis que neige et glace apparaissent dans les régions polaires.
Quelques organismes caractéristiques :
Organismes marins :
Unicellulaires : Diversification et expansion des Coccolithophoridés
C'est au Jurassique que ce groupe d'Haptophytes devient une composante notable du registre fossile, un groupe diversifié et un acteur sédimentaire important.
Invertébrés : Ammonites et Bélemnites
|
Ces deux types de Céphalopodes sont abondants dans les mers jurassiques, comme en atteste ce calcaire Jurassique supérieur (160 Ma). En noir, les rostres de Bélemnites, partie terminale de la coquille interne de l'animal (homologue de "l'os" de la seiche ou de la "plume" du calmar actuel).
Image : National Museum of Natural History, New York. échelle 1/2 |
|
|
|
Ci-contre, reconstitution d'une Belemnite. Image : BBC, Walking with Dinosaurs |
|
|


Vertébrés :
Reptiles : Ichtyosaures, Plésiosaures et Pliosaures.
Ces trois groupes de reptiles, entièrement revenus à la vie aquatique, dominent le réseau trophique marin, avec les crocodiles.
|
Les Ichtyosaures sont désormais parfaitement adaptés à la vie au large, et à une nage rapide comme celle des thons actuels, avec un corps ramassé et fuselé aux vertèbres courtes et aplaties assurant une grande rigidité à la colonne vertébrale. Certaines formes s'adaptent à la plongée à grande profondeur, avec des yeux de très grande taille en proportion du reste du corps (comme chez Ophtalmosaurus, ci-contre). |
|

|
Image : S. J. Gould (dir.), Le livre de la vie, Seuil |
Enfin, ils acquièrent une reproduction ovovivipare. Un fossile exceptionnel (ci-contre) montre une femelle morte au cours de l'accouchement. La mère et le jeune (sur la droite, sortant de la mère la queue en avant) ont été tout deux fossilisés. |

|
|
Le groupe des Plésiosaures se diversifie en deux branches, les Plésiosaures au sens strict, à long cou, et les Pliosaures, au cou plus court. Certains atteignent des tailles très importantes. Le plus grand genre, un Pliosaure, Liopleurodon, (ci-dessous) atteint 12 à 15 mètres de long. Reconstitutions: site internet de la BBC Walking with Dinosaurs (série de vulgarisation scientifique, diffusée en version française sous le titre Sur la terre des dinosaures) |

Organismes terrestres :
Les plantes du Jurassique :
La végétation du Jurassique est dominée par les Gymnospermes, ensemble paraphylétique comprenant Conifères, Cycadophytes et Ginkgophytes, et par les deux groupes d'Anthophytes les plus primitifs, les Benettitales, (ou Cycadeoidophytes, entièrement éteintes) et les Gnetophytes.
Feuilles d'un Ginkgo actuel. 
Cependant, le plus ancien fossile connu de plante à carpelle (structure caractéristique des Angiospermes, dernier taxon d'Anthophytes) est daté de 140 Ma, donc du Jurassique récent. Ce fossile trouvé en Chine, Archaefructus liaoningensis, est constitué d'un fragment de tige portant des fruits. On n'en connaît pas le pollen.
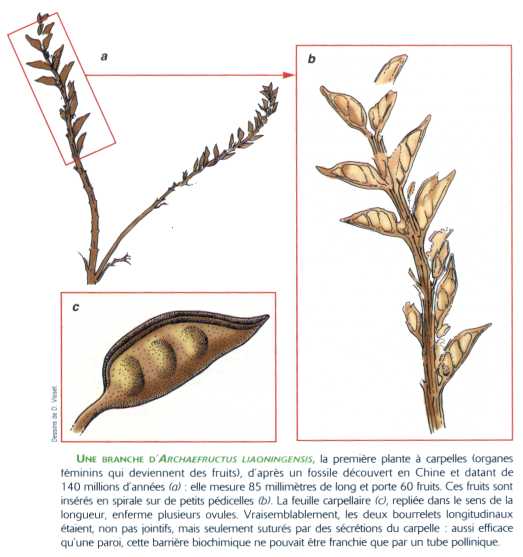
Document: Dossier spécial Pour la Science, De la graine à la plante, janvier 2000
La domination des Dinosaures :
Le groupe des Dinosauria profite de l'extinction de la fin du Trias pour se diversifier, et devenir le groupe dominant de vertébrés terrestres. Si ses premiers représentants triasiques étaient surtout des carnivores bipèdes de taille moyenne, les suivants sont de toute tailles, carnivores comme herbivores, bipèdes ou quadrupèdes.
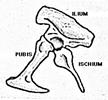
On subdivise cette classe en deux ensembles, selon l'anatomie de la ceinture pelvienne, les Saurischiens ("à bassin de reptile", anatomie primitive que l'on rencontre chez tous les Archosaures) et les Ornitischiens ("à bassin d'oiseau", disposition "dérivée"). Les deux groupes se seraient séparés dès le Trias récent.
|
|
Bassin d'Ornitischien |
|
L'ischion (ischium) pointe vers l'arrière et le bas. La tête de l'animal se trouve donc sur la gauche des figures. |
|
 Bassin de Saurischien
Bassin de Saurischien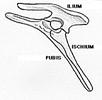
Les Saurischiens jurassiques les plus représentatifs appartiennent au groupe des Sauropodomorphes, qui comprend lui-même les Prosauropodes, (apparus à la fin du Trias) et les Sauropodes, qui en dérivent, au début du Jurassique. Ces derniers, les plus connus, sont des herbivores quadrupèdes, à long cou, longue queue et crâne relativement petit. La plupart sont de très grande taille. Aucun animal terrestre n'atteindra plus de telles dimensions.
Anecdote : ces tailles, et ces poids, gigantesques, ont conduit les paléontologues du XIXième et du début du XXième siècle à postuler que ces animaux vivaient immergés dans des marécages, la poussée d'Archimède soutenant ainsi leur masse. De nombreuses illustrations les ont représentés ainsi (par exemple le film Fantasia, de Walt Disney). Depuis, de meilleures estimations de la résistance mécanique des os, de nouvelles reconstitutions anatomiques des squelettes, et l'étude plus approfondie des milieux sédimentaires d'où provenaient les fossiles, ont établi leur mode de vie entièrement terrestre, probablement en troupeaux.
Notons encore que ces animaux, avec leur faible volume crânien comparativement à la taille de leur corps, sont aussi à l'origine du mythe de la Ç stupidité È des Dinosaures.
Parmi les plus connus :
- Diplodocus (Amérique du Nord,~27 m de long, ~20 tonnes ; un fossile récent présentant la trace de la peau a montré que cet animal portait sur le dos une crête d'épines ; d'autres sauropodes partageaient peut-être ce caractère).
- Brachiosaurus (Amérique du Nord, Portugal et Algérie, ~22 à 30 m ; son poids aurait pu atteindre jusqu'à 80 tonnes)
- ou encore Mamenchisaurus (~26 mètres, Chine et Mongolie ; il se distingue par le plus long cou qu'ait jamais eu un animal terrestre).
|
|
|
|
Squelette de Diplodocus, Musée Carnegie, New York. Les visiteurs donnent l'échelle. |
Reconstitution de deux Mamenchisaurus |
|
|
|
|
Crâne de Brachiosaurus |
Reconstitution de Brachiosaurus |




Parmi les Saurischiens, on peut encore mentionner le groupe des Théropodes, Dinosaures bipèdes, et parmi ceux-ci les Carnosaures, grands carnivores, dont un représentant typique au Jurassique est Allosaurus, d'une dizaine de mètres de long.
|
Les Ornitischiens sont, quant à eux, représentés au Jurassique par des Dinosaures herbivores "à armures" ou "à plaques", l'Ordre des Thyreophores (Stegosauria + Ankylosauria). Tous portent des plaques osseuses sur le dos. Le plus représentatif au Jurassique est Stegosaurus (Stegosauria, Amérique du Nord, Jurassique récent, ~9 m, 2 tonnes). Il portait des plaques triangulaires dont la fonction exacte n'est toujours pas élucidée : protection, structure d'ornementation sexuelle ou de régulation thermique ?. L'extrémité de la queue présentait quatre pointes osseuses, sans doute utilisées par l'animal pour se défendre. |
Photo : Brendan Smith |

Les Vertébrés volants : Ptérosaures et Dinosaures à plumes :
Le groupe des Ptérosaures est considéré comme phylogénétiquement proche des Dinosaures, même si on ne connaît pour l'instant aucune forme "intermédiaire".
|
Ils sont présents dès la fin du Trias avec un premier groupe, les Rhamphorhynchoidés, dont les représentants sont de petite taille, mais déjà parfaitement capables d'un vol battu, et possèdent une queue, qui faisait probablement office de gouvernail. On les rencontre encore au Jurassique, avec des formes comme Dimorphodon (Jurassique ancien), Pterodactylus, ou Rhamphorhynchus (Jurassique récent, ci-contre). Ils s'éteignent à la fin du Jurassique. |
|
|
|
Enfin, l'analyse de traces retrouvées sur des dalles de calcaire, notamment en France, à Crayssac, a permis de montrer que ces animaux se déplaçaient au sol en prenant appui sur leur quatre membres, à l'instar des chauves-souris actuelles (reconstitution ci-contre). |


C'est aussi à partir des terrains du Jurassique moyen et supérieur que l'on trouve des représentants du deuxième ensemble de Ptérosaures, les Pterodactyoidés, des animaux de plus grande envergure, probablement capable de voler sur de grandes distances, sans queue, mais fréquemment pourvus d'une crête osseuse. Ils s'épanouiront au Crétacé.
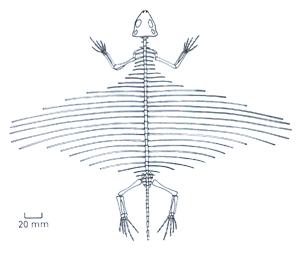
Des Dinosaures à plumes, capables au moins d'un vol plané, sont déjà présent au Jurassique récent, puisque les fossiles du premier et du plus célèbre d'entre eux, Archaeopteryx (ci-dessous), ont été retrouvés dans des calcaires Jurassique supérieur. En 2004,l L'étude fine de la minuscule boite crânienne (par une technique de tomographie aux rayons X) de l'un des sept spécimens connus d'Archaeopteryx a permis de montrer que son cerveau (quoique plus petit que celui d'un oiseau de même poids) avait déjà une organisation semblable à celle des oiseaux actuels, et qu'en particulier les aires cérébrales permettant le contrôle du vol y étaient déjà bien développées (Alonso et al., Nature 430, 5 aout 2004). Autrement dit, l'Archaeopteryx était doté d'un certains nombre des caractères qui permettent aujourd'hui aux oiseaux un vol efficace.
|
|
|


Des fossiles de dinosaures incapables de voler, eux, mais néanmoins pourvus de plumes, ont été depuis exhumés dans des terrains jurassiques et crétacés de Chine. Certains (en particulier Microraptor gui, du Crétacé ancien, décrit en 2003, et qui présentait de longues plumes sur les quatre membres) devaient cependant planer à la manière des écureuils volants actuels. Incapables de vol battu, (même s'ils sont plus récents qu'Archaeopteryx) ils ne sont pas encore considérés comme des Oiseaux, mais toujours comme des Dinosaures. Ces fossiles permettent maintenant de considérer les Oiseaux comme issus des Dinosaures Saurichiens Théropodes.
CRETACE (135-65 Ma)
L'environnement :
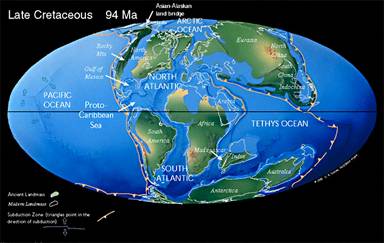
La Téthys est désormais raccordée à l'Atlantique, qui poursuit son ouverture vers le Nord. Le Gondwana est maintenant éclaté, et l'Inde, en particulier, commence à migrer vers le Nord. Cette fragmentation augmente la surface des mers épicontinentales. Le Crétacé présente une transgression généralisée. Dans ces mers prolifèrent les algues, et en particulier les Coccolithophoridés, à l'origine des vastes dépôts de craie qui valent son nom à cette période.
Le début du Crétacé n'est marqué par aucun bouleversement réel, dans le climat, la faune, ou la flore, par rapport au Jurassique.
Quelques organismes caractéristiques :
Organismes aquatiques :
Les Calpionnelles de la Téthys :
C'est à la fin du Jurassique, dans les séries téthysiennes calcaires, que l'on voit apparaître les premières Calpionnelles. Ces fossiles calcaires, en forme de petite amphore, sont aujourd'hui interprétés comme des loges d'unicellulaires Ciliés planctoniques. Ils présentent une importante et brève radiation évolutive au Crétacé ancien, et constituent par conséquent d'excellents fossiles stratigraphiques.
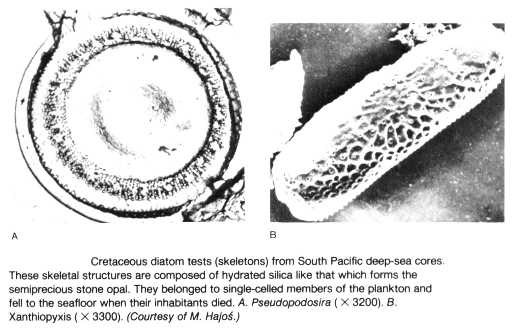
Les Diatomées marines :
Les premiers fossiles indubitablement attribués à des Diatomées datent du Crétacé ancien. Ce sont des formes marines. Les Diatomées d'eau douce n'apparaissent que beaucoup plus tard, au Miocène.
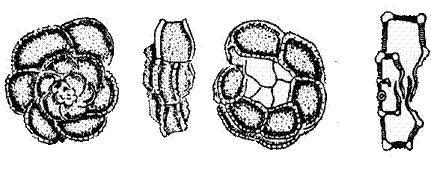
Foraminifères : les Globotruncana
Ces Foraminifères planctoniques à test hyalin, reconnaissables à leur carène latérale, se rencontrent fréquemment dans les sédiments du Crétacé supérieur, pour lequel ils permettent de faire des subdivisions biostratigraphiques fines.
. 
Mollusques :
Céphalopodes
Ammonites et Bélemnites sont toujours présentes.
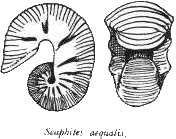
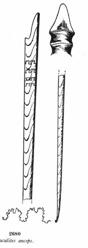
Cependant, l'adaptation des Ammonites aux milieux peu profonds des mers épicontinentales se traduit par l'apparition de nombreuses formes plus ou moins déroulées, comme (ci-dessous) Turrilites, Scaphites ou Baculites, à la coquille parfaitement droite.
Remarque : On peut visuellement rattacher ces fossiles aux Ammonites par la présence des sutures persillées, et dans le cas de Turrilites, en particulier, on peut distinguer cet animal d'un Gastéropodes (une Turitelle, par exemple) en remarquant que chaque tour conserve une symétrie bilatérale.
|
|
|
|
|
Turrilites
|
Scaphites
|
Baculites
|
Bivalves : les Rudistes
Les Rudistes représentent un groupe de Mollusques Lamellibranches, aussi appelés les Pachyodontes ("dents épaisses"). Ils sont caractérisés par une charnière particulière, composée d'épaisses dents (qui leur valent leur nom) et de profondes fossettes, ainsi que par une coquille fortement inéquivalve, et un mode de vie benthique, fixé par l'une des valves (fréquemment la valve droite).
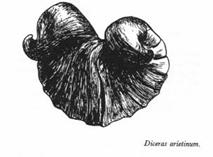
Les Rudistes les plus anciens, du Jurassique supérieur, comme Diceras, ci-dessous, montrent encore deux valves presque identiques.
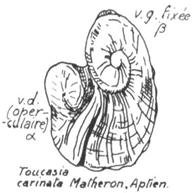
Les suivants ont un caractère inéquivalve beaucoup plus accentué (Requiena, ci-dessous), et, chez les Hippurites, la valve gauche se réduit à un opercule plat. la coquille de ces animaux ne s'ouvrait plus en charnière, mais par un soulèvement vertical de l'opercule.
|
|
|
|
|
Toucasia carinata Crétacé ancien |
Requiena ammonia Crétacé ancien |
Hippurites Crétacé récent |


Vertébrés : Reptiles marins.
|
Ichtyosaures et Plésiosaures sont toujours présents parmi les grands prédateurs marins. S'y ajoute au Crétacé le groupe des Mosasaures. (Ne pas confondre avec les Mésosaures du Permien. Le nom Mosasaure signifie "lézard de la Meuse", les premiers restes de ces animaux ayant été trouvé au Pays-Bas, près de ce fleuve). Aujourd'hui considérés comme apparentés aux Lézards, ces grands prédateurs, probablement piscivores, présentaient un corps sans cou, de 3 à 10m de long, et nageaient au moyen de leur queue et de membres modifiées en palettes natatoires.
Ci-contre : Reconstitution d'un Mosasaure. Dessin : J-M. Mazin, Dossier Pour la Science, La valse des espèces.
|
|

Organismes terrestres :
Végétaux : l'expansion des Angiospermes
Apparues au Jurassique, les Angiospermes se développent au Crétacé moyen, prenant le pas sur les Conifères alors que les Benettitales s'éteignent et que Cycadales et Gingkoales déclinent.
Arthropodes : Les Insectes
Le Crétacé représente un épisode de diversification évolutive des Insectes, qui auraient atteint alors une diversité comparable à l'actuelle. Ce succès paraît lié à l'expansion des Angiospermes. Insectes et plantes à fleurs auraient co-évolué, les premiers devenant les pollinisateurs principaux des secondes. C'est également dans les terrains Crétacé que l'on a trouvé les premiers Hyménoptères sociaux. Au Crétacé récent, toutes les familles actuelles d'Insectes sont déjà présentes.
Dans le même temps, cependant, à la limite Cénomanien-Turonien, plusieurs lignées anciennes, surtout d'eau douce, disparaissent.
Pour plus de détail, on pourra se reporter à l'article d'André Nel, Les insectes : un succès de l'évolution, dans Pour la Science, Mars 2002.
Vertébrés : Dinosaures et apparentés, toujours.
La faune de Vertébrés reste dominée par les Dinosaures, alors que les airs sont maintenant occupés par les grands Ptérosaures mais aussi par les Oiseaux.
Les formes de Dinosaures les plus connues du "grand public" sont d'âge Crétacé. On en détaille ci-après quelques-unes.
Ornitischiens :
Parmi ce groupe se développent alors plusieurs branches aux formes caractéristiques.
- A côté des Stégosauridés apparaissent les Ankylosauridés, groupe de dinosaures "cuirassés" au corps couvert de plaques osseuses défensives.

Euoplocephalus, ~6 m, l'un des Ankylosauridés les mieux connus, Crétacé supérieur d'Amérique du Nord.
Image : site Internet The Tree of Life
- Le groupe des Ornithopodes devient, au Crétacé, l'ensemble le plus diversifié des Ornitischiens, avec environ 80 espèces.
Parmi ceux-ci, les Hadrosaures, ou "dinosaures à bec de canard", développent une morphologie des mâchoires caractéristique : un museau en bec plat, édenté à l'avant, mais pourvu en arrière d'une batterie de dents aplaties. La zone avant permettait d'attraper et de découper efficacement les plantes dures, alors que la batterie dentaire conférait une grande surface de broyage.
|
D'autre part, certaines formes (comme ci-contre, Tsintaosaurus) présentent sur le crâne une crête, ou une expansion osseuse creuse, partant de la mâchoire supérieure, dont la fonction reste mal comprise. Hormis une utilisation comme signal visuel, des reconstitutions des flux d'air dans ces structures suggèrent qu'elles permettaient à ces animaux d'émettre des sons amplifiés.
Image : S.J. Gould, Le livre de la vie, Seuil. |
|

C'est également aux Ornithopodes qu'appartient le groupe des Iguanodontidés, et son plus fameux représentant, Iguanodon, premier dinosaure découvert, et décrit comme tel, au XIXième siècle.
|
Reconstitution d'un Iguanodon : Source :site Internet de la BBC, Walking with Dinosaurs. Taille : ~ 4 tonnes, 6 mètres |
|

- Les Pachycephalosaures ("tête épaisse") présentent, quant à eux, un crâne aux os fortement épaissis. On suppose que cette structure leur servait lors de combats entre mâles, à l'occasion des parades nuptiales, selon le même comportement que les Cerfs actuels.
- Enfin, le groupe des Cératopsiens, (dinosaures à collerette et à cornes) dont les premiers représentants (Protoceratops) remontent au jurassique récent, se diversifient au Crétacé et atteignent de grandes tailles. Triceratops, ci-dessous, en est l'un des représentants les plus connus.
|
Protoceratops, ~400 kg, 1,8 m
|
Triceratops (Image : UCMP). ~ 8 mètres, 6 à 12 tonnes |


Saurischiens :
- Les Théropodes, carnivores bipèdes, présentent plusieurs formes de très grande taille, aux membres antérieurs très réduits, dont le fameux Tyrannosaurus rex d'Amérique du Nord (~6 tonnes, 14 mètres de long, 5 m de haut), ou des formes équivalentes sur les autres continents, comme Tarbosaurus, trouvé en Mongolie.
|
|
|
|
Ci-dessus, crâne de Tyrannosaurus rex, exposé au Muséum de Paléontologie de Berkeley. |
|
|
Ci-contre, Tarbosaurus bataar, squelettes exposés au Museum de Paléontologie de Berkeley. Devant, le crâne d'un Hadrosaure. On reconnaît le museau en bec de canard typique de ce groupe |


D'autres Théropodes gardent des tailles plus modestes, des membres antérieurs relativement longs, mais présentent néanmoins des adaptations qui en faisaient sérement des prédateurs très efficaces. Ainsi, les Dromaesauridés, comme Velociraptor (~1,8m et 45 kg, Mongolie), ou Deinonychus ("griffe terrible", Amérique du Nord, représenté ci-dessous) possédaient sur un doigt du membre postérieur une grande griffe en faucille. On leur attribue souvent aujourd'hui, à l'inverse des Tyrannosauridés, un comportement de chasse en groupe. Notons enfin que c'est ce dernier groupe que les études cladistiques positionnent actuellement en groupe-frère des Oiseaux.
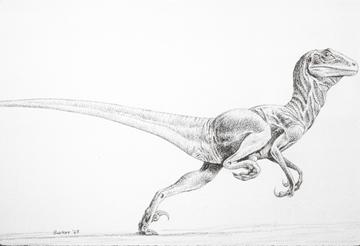
Oiseaux :
Au Crétacé, les vrais oiseaux sont déjà présents.
|
|
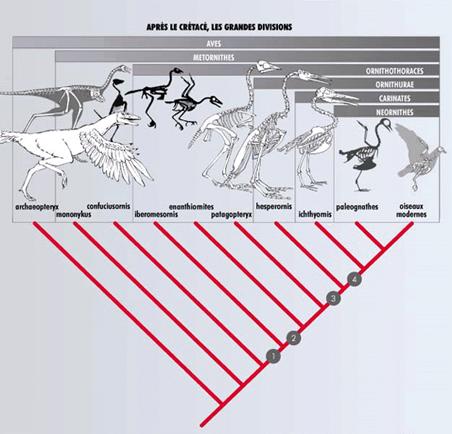
Jusqu'à récemment, aucun fossile ne permettait de relier Archaeopteryx aux fossiles d'oiseaux du Crétacé récent. De nouvelles découvertes, en particulier dans le Crétacé ancien de Chine, de Mongolie et d'Espagne, ont démontré l'existence de plusieurs lignées d'oiseaux au Crétacé. (cladogramme ci-contre. Source : La Recherche n° 317)
Leur mâchoire et leur crâne portent encore des dents. Certains, comme Hesperornis (ci-contre), sont revenus à un mode de vie pêcheur : incapables de voler, leurs pattes arrières sont très développées et sans doute palmées. |
Les Mammifères :
Les Mammifères sont déjà bien présents au Crétacé, et on en découvre de plus en plus de restes fossiles. Ils restent cependant de très petite taille. Les principaux appartiennent au groupe des Multituberculés, entièrement éteint. Se reporter à la feuille Paléocène.
Les sédiments Crétacé ont aussi fourni des fossiles attribués à des Monotrèmes (Crétacé inférieur d'Australie) et des Marsupiaux (Crétacé moyen d'Amérique du Nord).
La crise Crétacé / Tertiaire :
On ne détaillera pas ici les divers arguments avancés à propos des causes de cette extinction. Rappelons seulement qu'à l'heure actuelle, on l'attribue en premier lieu à l'impact d'un corps extraterrestre, et aux bouleversements climatiques qu'il aurait provoqué. On a retrouvé, par imagerie sismique, le cratère d'impact de cet objet au Yucatan (cratère de Chixculub). Les conséquences climatiques d'une gigantesque activité volcanique (également du même âge, et responsable de la mise en place des trapps du Deccan, en Inde) sont aussi invoqués comme facteur déclenchant de la crise.
Quel qu'en soit le responsable principal, la crise se traduit, pour la biosphère, par :
En domaine marin :
- La disparition de 16% des Familles, soit 45% des Genres et 76% des espèces (F. Lethiers, Evolution de la biosphère et événements géologiques, Gordon and Breach science Publishers).
-D'importants changements dans le microplancton (Foraminifères) et le nanoplancton (Coccolithophoridés), qui permettent une définition biostratigraphique très précise de la limite Crétacé/Tertiaire.
- En domaine marin toujours, Ammonites et Bélemnites disparaissent. Cependant, la diversité des Ammonites montre une baisse progressive avant la limite Cr/T, que l'on a mis en relation avec la régression marine concomitante. Il en va de même pour les Rudistes.
En domaine continental :
- La disparition de tous les animaux d'un poids supérieur à ~25 kg.
- Dinosauriens (à l'exception des Oiseaux), Ptérosauriens, Plésiosauriens disparaissent ainsi que, parmi les Lacertiliens, le groupe marin des Mosasaures (les Ichtyosauriens, eux, se sont éteints plus tôt, avant la fin du Crétacé). Cependant, parmi les Oiseaux, les formes "primitives" s'éteignent. Les oiseaux tertiaires appartiennent tous au groupe des Néornithes, les oiseaux actuels.
- Crocodiles et Tortues sont également affectés. Seuls les Amphibiens semblent passer cette période sans aucun dommage.
- Les Mammifères, tous de petite taille, sont peu ou pas affectés, à l'exception des Marsupiaux.
- Les plantes sont beaucoup moins touchées que les animaux, même si la limite Cr/T est marquée par un pic d'abondance de spores de fougères et de pollen d'Angiospermes, interprété comme la trace de grands incendies : destruction des plantes dont il reste les grains de pollen, puis recolonisation par les Fougères.
Epoque précédente / Retour en haut / Epoque suivante
PALEOGENE
1-PALEOCENE (65-53 Ma)
L'environnement :
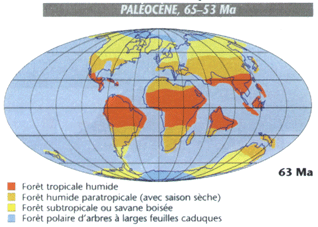
Le climat du Paléocène apparaît en moyenne plus froid que celui du Crétacé. Il reste néanmoins plus chaud que l'actuel, et surtout plus humide qu'auparavant, avec des précipitations plus fortes et plus également réparties dans l'année. De là le développement de forêts plus denses qu'au Crétacé en Amérique du nord, et de mangroves sur le continent australien par 65° de latitude sud.
Note :
Le Paléocène est marqué par un bref épisode particulièrement chaud, le Maximum Thermique du Paléocène Récent (Late Paleocene Thermal Maximum, LPTM) durant lequel les températures des hautes latitudes semblent avoir atteint les valeurs les plus élevées du Cénozoïque. Les causes de cet épisode sont encore mal comprises. Les enregistrements isotopiques du carbone 13 obtenus pour cette période ont conduit à proposer comme explication la déstabilisation des chlathrates des sédiments océaniques profonds (hydrates de gaz stables aux pressions et températures de l'océan profond), suite à des modifications de la circulation océanique. Ce phénomène aurait libéré dans l'atmosphère d'importantes quantités de méthane, gaz à très fort effet de serre.
Quelques organismes caractéristiques :
Organismes marins :
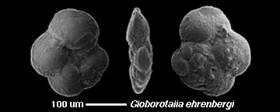
Foraminifères : les Globorotalia :
Proches des Globotruncana du Crétacé, ils apparaissent au Paléocène, et persistent jusqu'à aujourd'hui.
|
|
|

Organismes terrestres :
Les Oiseaux du Paléocène :
À partir du Paléocène, on retrouve deux grands clades d'Oiseaux, qui constituent ensemble le clade des Neornithes :
- Les Paléognathes ("mâchoire ancienne"), oiseaux "archaïques" aux ailes réduites, incapables de voler. Leurs représentants actuels sont essentiellement les Ratites (Kiwis, Autruches, Emeux et Casoars),
- Les Néognathes ("nouvelle mâchoire"), que l'on trouve dans le registre fossile essentiellement à partir de l'Eocène.
À la fin du Paléocène, puis à l'Eocène, en Amérique du Nord et en Europe, on rencontre de très grands Néognathes, comme Diatryma : incapables de voler, ils pourraient avoir été carnivores, chassant alors de la même manière que les grands dinosaures carnivores. Se reporter à la feuille Eocène.
Les Mammifères :
L'expansion des Mammifères intervient au Paléocène, suite à la disparition des Dinosaures, mais ils n'atteignent pas encore de très grandes tailles, d'autant que les forêts abondantes sont denses et fermées.
Remarques :
- Les restes fossiles de ces animaux ne sont souvent que des dents.
- Les groupes qui se développent alors n'ont, pour beaucoup, plus de représentants aujourd'hui.
La figure ci-dessous illustre bien la transition, rien qu'en terme d'abondance générique, entre faune crétacé à dinosaures et faune paléocène à mammifères.
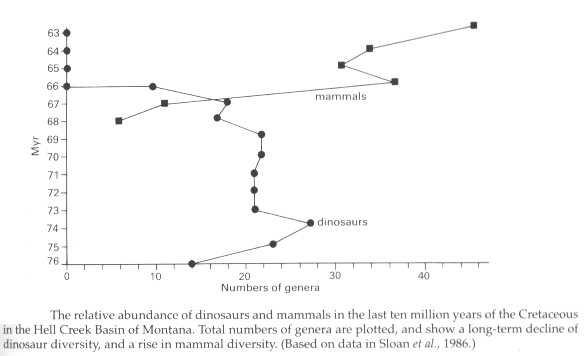
Figure : M. J. Benton, Vertebrate Paleontology, Blackwell Science.
Les Multituberculés :
Connu depuis le Jurassique moyen, ce groupe s'est développé au Crétacé récent, et est bien représenté au Paléocène.
La morphologie de ces animaux se rapprochait beaucoup de celle des Rongeurs actuels, et ils devaient occuper la même niche écologique : s'ils ne disposaient pas des incisives en ciseau à croissance continue qui caractérisent les Rongeurs (et ont assuré leur succès écologique), leur dentition présentait pourtant des spécialisations marquées: leurs incisives étaient allongées, les canines généralement absentes, et les dernières prémolaires inférieures formaient une large plaque aux crêtes, ou au tubercules, nombreux et allongés latéralement (d'où leur nom). Cette structure spécifique leur permettait de déchirer efficacement leur nourriture.
Certains, comme Ptilodus, ci-dessous, trouvé dans le Paléocène du Canada, devaient avoir un mode de vie grimpeur comme les écureuils actuels.
|
|
|
|
Ptilodus. Crâne et reconstitution. ~50 cm de long. Remarquez, sur le crâne, à gauche, la quatrième prémolaire inférieure, très grosse et munis de nombreuses crêtes allongées. Images : d'après Cox (1998), tirée du site Internet Paleocene Mammals, |
|
Les Monotrèmes et les Marsupiaux :
On ne dispose que de quelques fossiles fragmentaires de Monotrèmes avant le Miocène.
Les Marsupiaux paraissent fortement affectés par la crise Crétacé-Tertiaire. Leurs premiers fossiles, d'âge crétacé, proviennent surtout d'Amérique du Nord, d'où ils seraient passés en Amérique du Sud. C'est probablement au cours du Paléocène qu'ils migrent encore jusqu'en Australie, via l'Antarctique.
La plupart des Marsupiaux fossiles du Crétacé et du Paléocène rappellent les opposums actuels, et sont classés avec ces derniers dans le clade des Didelphoïdés.
Les Euthériens :
En Amérique du Nord, en Europe et en Asie, Les Placentaires montrent une rapide radiation évolutive au cours du Paléocène, avec :
- L'apparition et/ou la diversification de lignées aujourd'hui éteintes :
- des Ongulés primitifs, les Condylarthres et les Mésonychidés, se développent et, pour certains, s'adaptent à un régime carné. Les Mésonychidés comprendront les plus grands mammifères carnivores du Paléocène et de l'Eocène.
- A partir du Paléocène récent, on rencontre un autre groupe de mammifères carnivores, groupe-frère éteint des Carnivores actuels, les Créodontes.
- Plusieurs groupes d'herbivores.
- L'apparition, surtout à la fin du Paléocène, de plusieurs clades encore actuels, en particulier :
les Insectivores, les Carnivores "modernes", les Chiroptères (chauve-souris), les Primates, les Périssodactyles et les Rongeurs.
- Primates :
|
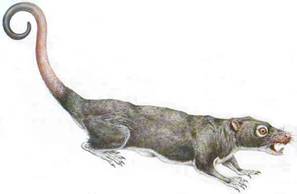
C'est dans le Crétacé et surtout le Paléocène inférieur d'Amérique du Nord que l'on a trouvé des restes d'un mammifère attribué au clade des primates, Purgatorius, de la taille d'un rat (ci-contre).
Image : d'après Cox (1998), site Internet Paleocene Mammals, www.paleocene-mammals.de
|
|

|
|
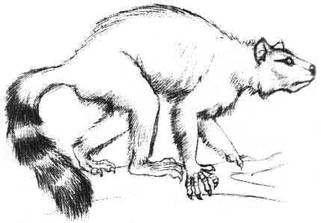
Au Paléocène récent apparaît le clade des Plésiadapiformes et les Plésiadapidés. Ci-contre, une reconstitution de Plesiadapis, qui donne son nom au groupe.
Images : d'après Cox (1998), site Internet Paleocene Mammals, www.paleocene-mammals.de
|
Une reconstitution :
|
Cette image donne une idée des forêts du début du Paléocène, dans l'actuel Wyoming ( Etats-Unis). Les grands arbres sont des séquoias. Les arbustes comprennent des théiers et des lauriers, mêlés à des fougères et à des prêles. Sur l'arbre, au centre, le multituberculé Ptilodus. Plus haut, en gris, un marsupial, Peradectes. Plus gros, à gauche, Chriacus, un omnivore, (représentant du groupe éteint des Arctocyonides, (une branche de l'ensemble des Condylarthes, un groupe peut-être hétérogène). Image : S. J. Gould, Le livre de la vie, Seuil |
|

PALEOGENE
2-EOCENE (53-34 Ma)
L'environnement :
|
|
|
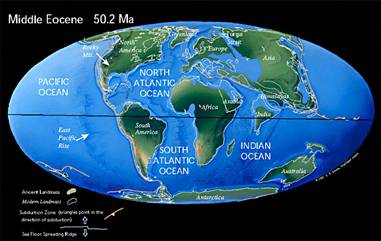
L'Eocène est une période longue, au cours de laquelle s'épanouissent de nombreux grands groupes, pour beaucoup encore représentés aujourd'hui, en particulier parmi les Vertébrés et surtout les Mammifères.
Le climat est toujours chaud, avec un faible gradient latitudinal.
Notons enfin que c'est vers entre -55 et -45 Ma que commence la collision de l'Inde et de l'Eurasie : les premiers signes de raccourcissement de la marge indienne datent d'environ -55 Ma, le ralentissement notable de la progression de l'Inde vers le Nord, suite au contact des deux croûtes continentales, de -45 Ma.
Quelques organismes caractéristiques :
Organismes marins :
Foraminifères :
Les Nummulitidés :
Les Nummulitidés sont un groupe de Foraminifères benthiques de grande taille, facilement reconnaissable en lame mince. le genre Nummulites est caractéristique des sédiments Eocène et Oligocène.
|
|
|
|
Nummulites dans un calcaire Lutétien (Eocène inférieur). Ci-dessus, coupe axiale ; Ci-contre, coupe équatoriale. |


Les Alvéolinidés :
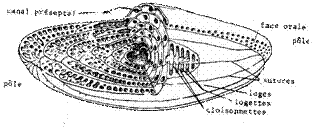
Présents du Crétacé récent à aujourd'hui, ces Foraminifères sont bien présents à l'Eocène, en particulier avec le genre-type Alveolina (coupe ci-dessous), qui donne son nom au groupe.
Echinides : les oursins irréguliers plats
En conséquence du développement des milieux de plages sableuses, on voit se développer à l'Eocène des oursins irréguliers fouisseurs très aplatis, les "dollars des sables" (sand dollars). Ils évoluent à partir d'espèces plus bombées.
Le retour à l'eau des Mammifères :
Les Siréniens :
Les plus anciens fossiles de ce taxon [groupe-frère des Proboscidiens (éléphants)] datent de l'Eocène ancien. Ils se diversifient au cours de l'Oligocène et du Miocène.
Les Cétacés :
Les premiers Cétacés vrais apparaissent à l'Eocène. Tous les fossiles découverts à ce jour proviennent du Pakistan et d'Egypte. Jusqu'à 2001, les origines et le plus proche parent taxinomique de ce groupe restaient énigmatiques. Les fossiles disponibles jusque-là rapprochaient les Cétacés des Mésonychidés, mammifères terrestres à sabot, eux-mêmes proches des Artiodactyles. La phylogénie moléculaire sur les groupes actuels plaçait plutôt les Cétacés en groupe-frère des Artiodactyles terrestres.
La découverte et la description, au cours des années 2000-2001, de nouveaux fossiles pakistanais, ont permis de réconcilier ces deux conceptions, en apportant des arguments morphologiques concordant avec les données génétiques pour faire des Cétacés une branche des Artiodactyles, les Mésonychidés passant alors en groupe-frère de l'ensemble (Cétacés + autres Artiodactyles terrestres).
Ce n'est qu'après l'Eocène que les Cétacés se diversifient en deux branches : les Odontocètes , les baleines à dents, (dauphins et orques) et les Mysticètes, baleines à fanons (baleine grise, rorqual [= baleine bleue], et cachalot [bien que lui ait conservé des dents])
L'évolution des Cétacés, un exemple de complémentarité entre biologie moléculaire et paléontologie.
Les études paléontologiques se basaient pour l'essentiel, avant 2001, sur le crâne d'un Cétacé archaïque, Pakicetus, et sur le squelette incomplet d'un grand Cétacé déjà entièrement aquatique, aux membres postérieurs atrophiés, Basilosaurus (ci-dessous).

Basilosaurus, étymologiquement "lézard-roi", car initialement décrit comme un reptile. -47 à -42 Ma, découvert en Egypte. ~20 mètres de long.
|
Ces fossiles apparentaient les Cétacés aux Mésonychidés, mammifères terrestres à sabot, eux-mêmes groupe-frère putatif des Artiodactyles, selon le schéma : |
Artiodactyles + (Mésonychidés + Cétacés)
|
|
Les nouveaux fossiles exhumés au Pakistan et décrits en 2001 conduisent à rattacher les Cétacés aux Artiodactyles, replaçant les Mésonychidés en groupe externe du clade (Cétacés + Artiodactyles): |
Mésonychidés + (Cétacés + autres Artiodactyles)
Conclusion qui rejoint celles de la génétique moléculaire. |
L'une des principales données paléontologiques qui a permis de rattacher les Cétacés aux autres Artiodactyles provient de la découverte, sur ces nouveaux fossiles, de l'astragale (os de la cheville entièrement atrophié sur les Cétacés actuels) : cet os a, chez les Artiodactyles, une forme caractéristique, avec deux extrémités en "poulie". Or, l'astragale des fossiles possédaient également ce caractère, qui par contre est absent chez les Mésonychidés. il s'agit donc d'une apomorphie de l'ensemble (Cétacés+Artiodactyles).
De plus, ces spécimens permettent de concevoir la série de transitions qui a fait passer d'Artiodactyles terrestres coureurs, (comme Diacodexis), à des Cétacés entièrement aquatiques, aux membres postérieurs réduits, comme Basilosaurus ou Dorudon, puis les baleines actuelles.
|
|
|
|
Ci-dessus : Rhodocetus, -46,5 Ma, une forme proche d'Ambulocetus, Pakistan. Dans les deux cas, ces Cétacés archaïques devaient mener une vie semi-aquatique, à l'instar des Phoques actuels, et se propulsaient dans l'eau par battements de leurs pattes arrières. Source : Gingerich et al. (2001), Origin of whales from early artiodactyls: hands and feet of Eocene Protocetidae from Pakistan. Science 293: 2239-2242 |
Une série de squelettes allant de l'Artiodactyle primitif Diacodexis jusqu'à la Baleine actuelle, en passant par quelques-uns des fossiles découverts au Pakistan : Pakicetus, (-52 Ma), Ambulocetus, ƒocène moyen, Dorudon, ƒocène récent. Source : De Muizon. (2001), Walking with Whales, Nature 413: 259-260
|

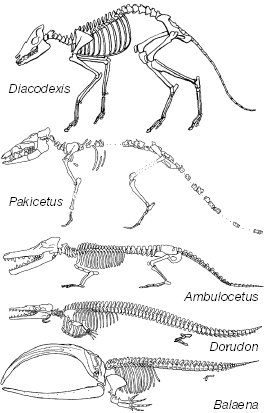
Organismes terrestres :
Oiseaux :
|
Comme mentionné dans la section Paléocène, les Néognathes comprennent en particulier, à cette époque, des oiseaux de grande taille (>2m), peut-être carnivores. Ci-contre, une reconstitution de l'un d'entre eux, Gastornis. Beaucoup d'ouvrages mentionnent Diatryma, un autre représentant de ces animaux (ci-dessous).
|
Images : Ci-dessus: Site web de la BBC, Walking with Beasts. Ci-contre : Encyclopaedia Universalis. |
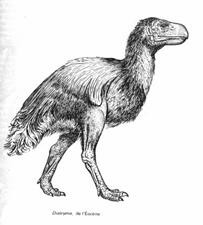

Mammifères :
Au sein des nombreux groupes de Mammifères terrestres présents à l'Eocène, on trouve certains des plus spécialisés, et la plupart des grands groupes actuels. Parmi ces derniers, on peut citer :
- Les Proboscidiens : Ce groupe d'animaux "à trompe (proboscis)" apparaît lui aussi à l'Eocène. La trompe elle-même n'est évidemment pas un organe fossilisable. L'attribution d'un fossile à ce taxon repose donc sur d'autres caractères. La présence effective d'une trompe et sa forme peuvent néanmoins se déduire de l'étude de la morphologie du crâne.
|
Ci-contre, Moeritherium, petit proboscidien (~0,7m au garrot) trouvé d'abord en Egypte, puis en Afrique du Nord et de l'Ouest. fin de l'Eocène, début de l'Oligocène. Image : Site web de la BBC, Walking with Beasts |
|

- Les Artiodactyles : avec par exemple Diacodexis, mentionné plus haut.
- Les Périssodactyles : ce groupe se diversifie énormément durant cette période, et y atteint son apogée. Ses clades principaux sont alors tous représentés de par le monde: Rhinocérotidés, Equidés, Tapiridés, Brontothérioidés (encore appelé Titanothérioidés) et Chalicothérioidés (exclusivement européens au départ, ces animaux se dispersent à l'Eocène moyen en Asie et en Amérique du Nord). Les deux derniers groupes sont entièrement fossiles, mais important à l'Eocène.
Parmi ces Périssodactyles éocènes, signalons les animaux suivants :
|
|
Bronthothérioidés : Groupe-frère des autres clades de Périssodactyles, il a compris, à la fin de l'Eocène et à l'Oligocène, des animaux énormes, portant des cornes nasales compliquées. Ci-contre, par exemple, Embolotherium (~2,5m au garot) Remarque : leurs cornes étaient constituées d'os, donc fossilisables, et non de kératine comme chez les Rhinocérotidés. Image : BBC, Walking with Beasts |
|
|
Equidés : Hyrachotherium (à droite), trouvé en Amérique du Nord et en Eurasie, est l'un des premiers représentants de ce clade. De la taille d'un chien, il possède encore quatre doigts bien développés à la patte avant et trois à l'arrière.(ci-contre). Il s'éteint en Europe au cours de l'Eocène |
|
Image : Science & Vie, hors série. Décembre 2000
|
|
Image : Science & Vie, hors série. Décembre 2000 |
On peut encore mentionner : - Propalaeotherium, un second équidé éocène, d'apparence très proche de celle d'Hyrachotherium. On en a retrouvé des fossiles très bien conservés dans l'exceptionnel gisement de Messel, en Allemagne. - Palaeotherium (ci-contre), à l'allure très proche de celle d'un tapir qui apparaît en Asie avant de coloniser l'Europe, à l'Eocène moyen et récent. |
|


Rhinocérotidés et Tapiridés sont représentés par des formes de taille modeste, extérieurement assez semblables aux Equidés contemporains.
Notons enfin un groupe de périssodactyles aujourd'hui éteint, les Lophiodontidés, avec pour représentant typique Lophiodon, déjà décrit par Cuvier en 1822. Les taxons de ce groupe, associés aux Palaeothéridés ci-dessus (Palaeotherium entre autres), sont caractéristiques de la faune endémique que l'on trouve dans les gisements de l'Eocène récent d'Europe, en particulier en France (dans les calcaires Lutétien du bassin de Paris, par exemple).
- Les Chiroptères : la plus ancienne chauve-souris connue est datée de -55 Ma (fin du Paléocène).
- Les Rongeurs :
On suppose que la disparition des Multituberculés, qui s'éteignent à la fin de l'Eocène, serait liée à leur compétition avec les Rongeurs pour les mêmes niches écologiques. Les incisives hypsodontes et en ciseau des Rongeurs constitueraient une "adaptation-clé", qui aurait assuré leur réussite au détriment des Multituberculés.
- Les premiers Lagomorphes, les actuels lapins et lièvres, datent également de l'Eocène
- Les Primates :
Les primates de l'Eocène, que l'on trouve en Asie, en Afrique et en Amérique du Nord présentent déjà la plupart des caractères typiques du groupe : pied préhensile, yeux surmontés d'une barre post-orbitaire, tournés vers l'avant et permettant donc la vision binoculaire, ongles au lieu de griffe·On ne connaît donc pas l'histoire de l'acquisition de ces caractères.
La majorité des Primates éocènes, en particulier ceux d'Eurasie et d'Amérique du Nord, appartiennent à plusieurs groupes, les plus importants étant :
- les Notharctidés (Amérique du Nord, Europe et Asie)
- et les Omomyidés (Amérique du Nord).
Ces taxons disparaissent après l'Eocène, et tous les primates suivants proviennent d'autres groupes, asiatiques et africains, eux aussi présents dès l'Eocène. Tous les primates actuels descendent des taxons africains.
Parmi les groupes éteints, indiquons encore :
- Les Mésonychidés :
|
Ce groupe de quadrupèdes ongulés, d'origine Paléocène, s'est diversifié à l'Eocène, et comprends entre autres, à la fin de cette période, un remarquable carnivore, découvert en Mongolie, Andrewsarchus : Sa tête mesurait plus d'un mètre de long et son poids est estimé à près d'une tonne. |
Image : BBC, Walking with Beasts |

La crise Eocène / Oligocène :
La limite Eocène-Oligocène, il y a ~34 Ma, est marquée sur les terres émergées par un remaniement faunique très important, qui lui a valu, d'abord pour les terrains d'Europe, le nom de "Grande Coupure". Aujourd'hui, on constate qu'il s'agit, pour les faunes mammaliennes en particulier, d'un bouleversement global.
En Amérique du Nord, la faune terrestre subit plusieurs épisodes d'extinctions successifs à la fin de l'Eocène.
En Europe, suite à une importante régression marine à la fin de l'Eocène, une mer intérieure, qui formait une barrière entre l'Asie et l'île (ou l'ensemble d'îles) qu'était alors l'Europe, disparaît. Les mammifères asiatiques peuvent donc passer en Europe. Mieux adaptés aux nouvelles conditions climatiques, et aux changements de végétation et de régime alimentaire qu'elles entraînent, ils y remplacent bon nombre d'espèces indigènes.
D'autres modifications fauniques ont maintenant été mises en évidence à cette même époque en Asie centrale et en Asie du Sud-Est.
Dans les océans, on enregistre également un important épisode d'extinction. Par exemple, pour les Mollusques, seulement 10% des espèces éocènes sont retrouvées dans les terrains oligocènes.
Tous ces bouleversements sont, le plus souvent, mis en relation avec les changements climatiques qui interviennent à la fin de l'Eocène :
- Les mesures isotopiques de l'oxygène (18O) enregistrées par les coquilles des invertébrés marins (Foraminifères p.ex.) indiquent une diminution de la température moyenne des eaux du pacifique, de 22°C à 12°C.
- Parmi les organismes, marins en particulier, les plus affectés par la crise appartiennent à des espèces acclimatés aux eaux chaudes.
|
- Sur la terre ferme, en Amérique du Nord par exemple, on constate un changement floristique, avec la diminution des plantes à feuilles large et entières, indicatrices de climats chauds et relativement humides, au profit d'espèces aux feuilles plus découpées, comme le montre la figure ci-contre (de S. M. Stanley, Earth and Life through time) Notes : "entire-margined species" : espèces à feuilles entières
- Le climat général devient donc plus froid, plus aride sur les continents aux basses latitudes, et plus saisonniers, avec sans doute des hivers plus froid et un contraste été-hiver plus marqué. - La glace commence à s'installer sur l'Antarctique, ce qui entraîne le développement d'une psychrosphère dans les profondeurs de l'océan.
|
|
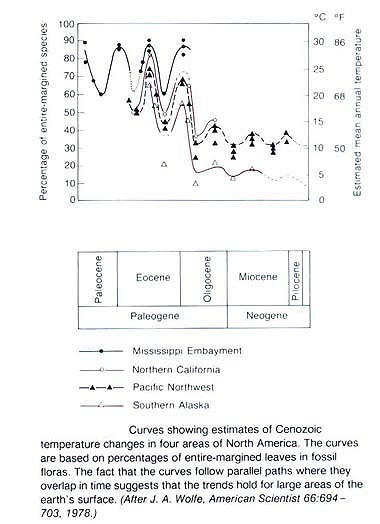
NEOGENE
1-OLIGOCENE (34-23 Ma)
L'environnement :
L'Oligocène est une période d'extension des forêts tempérées aux latitudes moyennes et élevées, mais la toundra reste limitée aux très hautes latitudes, en Antarctique. Une calotte glaciaire se développe sur ce continent, alors que le pôle Nord reste libre de glace. Le climat global est donc plus froid qu'auparavant, plus proche de l'actuel, sans cependant atteindre le gradient latitudinal de température et de végétation que l'on rencontre aujourd'hui.
Quelques organismes caractéristiques :
Organismes terrestres :
Mammifères marsupiaux :
L'Amérique du Sud, isolée pendant la plus grande partie de l'ére tertiaire, voit se développer une faune endémique, très peu apparentée à celles des autres continents, et dominée par les Marsupiaux, qui montrent souvent d'étonnantes convergences morphologiques avec les Euthériens des autres continents.
Mammifères euthériens :
Carnivores : Créodonta et Carnivora
La niche écologique des carnivores est partagée, à l'Oligocène, par plusieurs grands groupes. à côté du clade des Carnivora, qui contient aujourd'hui l'ensemble des carnivores actuels (Félidés, Canidés, Ursidés), on y trouve également des animaux placés dans leur groupe-frère, les Créodontes. ces derniers persisteront jusqu'au Miocène en Afrique et en Asie, mais s'éteignent à l'Oligocène en Amérique du Nord.
|
Parmi eux, le genre Hyaenodon, de Mongolie et d'Amérique du Nord, avec ci-contre la plus grande espèce, Hyaenodon gigas, environ 1,4 m au garrot. |
|

Les Artiodactyles :
L'Oligocène est une période de diversification des Artiodactyles. Les Suidés (porcs), les Camélidés (actuels chameaux et lamas) et les Ruminants (Bovidés, Ovidés, Cervidés) apparaissent.
|
|
On peut aussi mentionner le groupe des Entélodontidés, groupe-frère des Suidés, déjà présent à l'Eocène. Ces animaux, qui pour certaines espèces, pouvaient atteindre la taille d'une vache, se distinguaient par une tête très développée, aux dents et aux mâchoires fortes : on suppose donc qu'ils avaient un régime plus carné que les porcs actuels, et qu'ils se comportaient peut-être en charognard. Ci-contre, une reconstitution d'un Entelodonte. Image : BBC, Walking with Beasts |

Les Périssodactyles :
Les Périssodactyles sont toujours bien représentés à l'Oligocène.
En Europe sont arrivées des formes asiatiques de Rhinocérotidés ou de Chalicothérioïdés.
Les Equidés comprennent de nouvelles formes, comme Mesohippus en Amérique du Nord, qui n'a plus que trois doigts à la patte antérieure, et des prémolaires "molariformes", c'est-à-dire ressemblant à des molaires, autrement dit à la surface de broyage accrue, et qui permettent donc de consommer des feuilles plus coriaces qu'auparavant.
|
Les Chalicothérioidés de l'Oligocène d'Europe sont représentés par des animaux comme Chalicotherium (ci-contre), bien caractéristique de ce groupe d'herbivores. Les membres avant sont plus allongés que les pattes arrière, et l'animal se déplace à la manière des gorilles actuels, sur le dos des phalanges.
Image : BBC, Walking with Beasts |
|

Les Bronthothérioidés sont, eux aussi, nombreux, et, pour beaucoup, de grande taille (voir la section Eocène).
|
|
Les Rhinocérotidés sont abondants. C'est dans ce groupe que l'on trouve, dans l'Oligocène d'Asie, le plus grand mammifère terrestre connu, Paraceratherium (autrefois dénommé Baluchitherium, car découvert dans la province afghane du Baluchistan, ou encore Indricotherium). Cet énorme herbivore atteignait plus de 5 mètres au garrot, et 15 à 25 tonnes. Image : Pour la Science, Juillet 2000 |

Beaucoup d'autres formes de Rhinocérotidés, de taille plus modeste, avec ou sans corne, ont été exhumées dans les terrains oligocènes d'Eurasie et d'Amérique du Nord.
Remarque : Artiodactyles et Périssodactyles, un exemple de remplacement évolutif ?
Beaucoup d'ouvrages présentent encore l'Oligocène comme la période au cours de laquelle débute une radiation des Artiodactyles (apparition et/ou expansion de nouveaux groupes aujourd'hui importants, comme les Cervidés, les Suidés, les Hippopotames) et un déclin, en diversité, des Périssodactyles (disparition des Brontothéridés, raréfaction des Chalicothéridés et des Tapiridés).
Ce schéma a conduit à affirmer qu'il s'agissait là d'un cas de remplacement par compétition des Périssodactyles par les Artiodactyles, puisque les espèces des deux groupes ont des régimes alimentaires et des masses assez semblables, et partagent donc les mêmes niches écologiques. Certains travaux attribuaient le succès des Artiodactyles à une meilleure capacité à digérer leur nourriture.
Cette interprétation est aujourd'hui remise en question, suite à des études statistiques plus fines sur les taux d'apparition et de disparition d'espèces dans les deux clades. Il en ressort qu'ils auraient plutôt eu des histoires évolutives parallèles au cours du Tertiaire. De plus, l'efficacité supérieure de la digestion des Artiodactyles est également mise en doute.
Il reste vrai qu'actuellement, et depuis le Pliocène, les Artiodactyles sont bien diversifiés, et dominent la niche des herbivores de taille moyenne à grande, alors que les Périssodactyles sont réduit à trois groupes (Tapirs, Chevaux et Rhinocéros) et à quelques espèces seulement. Néanmoins, que ces deux histoires évolutives différentes soient le simple résultat d'une compétition écologique reste l'objet de débats. [Beaucoup plus que le cas similaire des Multituberculés face aux Rongeurs, au Paléocène]
Les Anthropoïdés (Simiens) :
L'apparition du clade des Anthropoïdea (= Simiens : Platyrrhiniens ["singes du Nouveau Monde"], et Catarhiniens [= Cercopithecoïdes et Hominoïdes, d'Afrique]) au sein des Primates s'est produite dès le début de l'Eocène (Le plus ancien fossile attribué à ce clade est Eosimias, récemment trouvé dans l'Eocène moyen (-40 Ma) de Chine).
|
Cependant, les fossiles les mieux étudiés datent de l'Oligocène, et sont classés parmi les Catarhiniens, dans le groupe des Propliopithécidés. On y place des fossiles découverts dans la région du Fayoum, en Egypte, comme Aegyptopithecus et Propliopithecus, des singes de 4 à 6 kg environ.
|
|

D'autre part, les singes Strepsirhiniens, ou "singes du nouveau Monde", aujourd'hui exclusivement américains, sont présents dès l'Oligocène en Amérique du Sud. Les groupes de primates Nord-américains de l'Eocène (Omomyidés) ont alors tous disparus, sans descendance, et les Strepsirhiniens Sud-américains proviennent donc de formes africaines, ce qu'indiquent aussi les analyses phylogénétiques. Or on comprend mal comment s'est effectué ce passage de l'Afrique à l'Amérique du Sud, dans la mesure où l'Atlantique Sud était déjà, à l'époque, largement ouvert.
NEOGENE
2-MIOCENE (23-5,3 Ma)
L'environnement :
|
|
|
|
Le Miocène est marqué par des changements climatiques et écologiques importants. Les courants marins sont fortement remaniés suite à deux évolutions tectoniques : -La séparation de l'Amérique et de l'Antarctique permet la mise en place d'un courant marin circum-antarctique. Il isole ce dernier continent, et diminue le mélanges des eaux polaires et des eaux tropicales. La calotte polaire antarctique peut alors s'étendre. - D'autre part, la plaque arabo-africaine se rattache à l'Asie, supprimant ainsi les échanges longitudinaux entre les reliques de la Téthys et l'océan Indien. La Téthys devient Méditerranée Le climat du début du Miocène est plus chaud que celui de l'Oligocène. Par la suite, vers -13,5 Ma, il devient rapidement plus froid et plus sec, ce que reflète, en particulier, l'apparition et l'expansion des prairies à graminées, aux latitudes moyennes de l'hémisphère Nord. |
|
Cette évolution est attribuée à la surrection de grandes chaînes de montagnes, en premier lieu l'Himalaya (voir l'encadré plus loin).
Enfin, la fin du Miocène montre une régression marine importante, qui aboutit à l'assèchement de la Méditerranée : c'est l'Evénement Messinien (du nom de l'étage stratigraphique qui enregistre ce bouleversement). Il entraîne la mise en place locale d'un climat plus froid et plus sec, et une expansion des savanes.
Modification climatique miocène, CO2 atmosphérique, et orogenèse himalayenne :
L'orogenèse himalayenne, qui débute vers 50 Ma, conna”t une phase d'accélération vers 10-8 Ma. Cet événement a eu d'importantes conséquences sur le système climatique, et peut-être même un impact direct sur les changements qui se produisent au Miocène. En effet, l'orogenèse aurait :
- d'une part, provoqué des modifications des circulations atmosphériques, et conduit à la mise en place du régime de mousson indienne.
-d'autre part, en augmentant la surface et le volume de roche soumis à l'érosion (au sens large), elles seraient à l'origine de la chute des teneurs atmosphériques en CO2 que l'on constate au cours de cet épisode. L'altération, avec la formation de minéraux argileux au détriment des tectosilicates (quartz, feldspaths), constitue en effet un "puit" de CO2. On peut en effet représenter l'altération des silicates en milieu aérien par les réactions générales :
CaSiO3 + 2 CO2 + 2 H2O --> Ca2+ + 2 HCO3- + H4SiO4
Puis : Ca2+ + 2HCO3- + H4SiO4 --> CaCO3 + SiO2 + CO2 + 2 H2O
D'où, au bilan, une perte d'une molécule de CO2 atmosphérique par molécule de silicate altéré.
Cette baisse du taux de CO2 atmosphérique, en diminuant l'effet de serre atmosphérique, aurait permis la diminution des températures.
Quelques organismes caractéristiques :
Organismes marins :
Unicellulaires : expansion des Diatomées d'eau douce
|
Les Diatomées ont colonisées les eaux douces depuis l'Eocène. Mais au Miocène, elles se diversifient considérablement, en particulier les Diatomées Pennales (Diatomées à test elliptique et aux pores disposés selon une symétrie bilatérale sur les valves, opposées aux diatomées Centrales, à frustule en disque). Les Diatomées atteignent alors leur diversité maximale.
Ci-contre, un exemple de Diatomée Pennale
|
|

Organismes terrestres :
Le développement des plantes herbacées, et de la photosynthèse en C4.
Plusieurs arguments paléontologiques indiquent que le refroidissement climatique du Miocène s'accompagne, pour la première fois, de l'expansion des prairies d'herbacées aux latitudes moyennes aux détriments des forêts.
Les données géochimiques, de leur côté, prouvent que ces plantes nouvelles ont acquis et utilisent la photosynthèse de type C4, comme le montre par exemple cet enregistrement isotopique dans des terrains d'Amérique du Nord (ci-dessous)
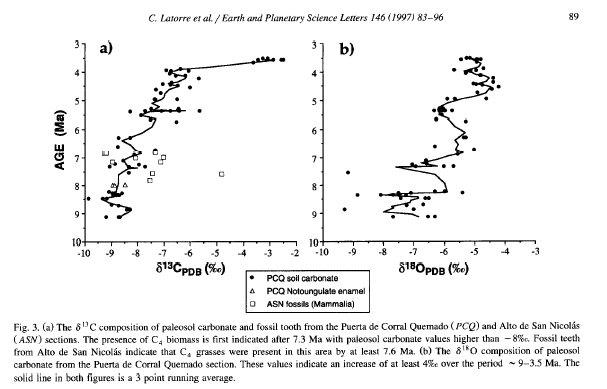
Cette figure présente à la fois :
- l'évolution des teneurs isotopiques en carbone 13, indice des proportions relative de plantes à photosynthèse en C3 (delta 13C bas, ~ -27o/oo) et en C4 (delta 13C plus élevé, ~ -13o/oo).
Les plantes en C3 ont des valeurs isotopiques du carbone de leur tissus basses, delta 13C ~ -27 o/oo, alors que la photosynthèse C4 conduit à des valeurs de delta 13C plus élevées, ~ -13 o/oo. Les carbonates des sols ou des dents des animaux enregistrent ces différences avec un décalage général des valeurs de ~ +14 o/oo.
-l'évolution des températures, reflétée par les teneurs en oxygène 18, d'autant plus élevées que la température locale est basse.
Remarque : Dans cette publication, les valeurs de delta 18O sont reportées par rapport au standard PDB, et non par rapport au standard SMOW. Autant que possible, on exprime désormais les valeurs isotopiques de l'oxygène relativement au SMOW, et l'on réserve le standard PDB aux valeurs isotopiques du carbone.
L'adaptation des herbivores aux plaines à herbacées :
Les Equidés : acquisition de l'hypsodontie et adaptation à la course
Ainsi que le montre la précédente figure, qui inclut des valeurs isotopiques de dents d'Equidés, ces animaux et d'autres herbivores ont progressivement inclus dans leur régime alimentaire les plantes en C4 de plus en plus présentes dans l'environnement.
Mais ces plantes, et plus généralement les herbacées, sont aussi plus siliceuse et donc plus abrasives que les feuilles tendres des arbres. Ce changement qualitatif de la nourriture explique l'apparition, chez les Equidés en particulier, de dents à l'émail épais, souvent très plissé pour former des crêtes coupantes, à couronne élevée, et hypsodontes (à croissance continue), toute ces adaptations permettant une meilleure résistance à l'usure. La figure qui suit, qui concerne encore les Equidés Nord américains, indique clairement l'évolution parallèle de la forme des dents et des teneurs isotopiques de l'alimentation, et le changement qui se produit vers -14 Ma.
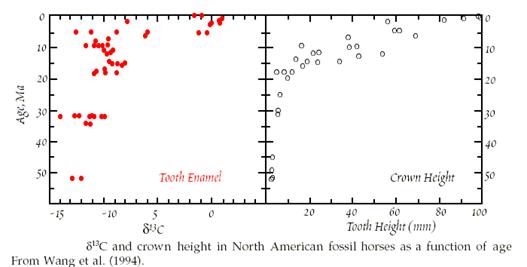
Enfin, ces nouveaux paysages ouverts se traduisent aussi, chez les Equidés entre autres, par une adaptation poussée à la course : par rapport aux premiers Equidés, les espèces miocène atteignent des tailles plus importantes, et se caractérisent par un allongement des membres et une réduction de plus en plus importante des doigts.
Les Hominidés : les plus vieux fossiles de ce clade à ce jour.
Sahelanthropus tchadensis, dit Toumaï, ~ -7 à -6 Ma :
|
Ce crâne, ainsi que des morceaux de la mâchoire et des dents, constituent les restes de ce que ses découvreurs décrivent comme le plus vieil Hominidé connu à ce jour, Kenyanthropus tchadensis, ou Toumai. Déterré en 2002 par une équipe franco-tchadienne, ce fossile bouleverse les connaissances paléoanthropologiques par deux particularités : -son âge, 7 Ma : Si Toumai appartient bien à la "lignée" humaine, le clade des Hominines, il réconcilierait les données fossiles avec les estimations de la génétique moléculaire, qui repoussent la divergence Homme-Chimpanzé à environ 8 Ma. |
|

- sa position géographique, au Tchad, donc très loin de l'Afrique de l'Est, d'où provenait jusque là la quasi-totalité des restes fossiles d'Hominidés. Il infirme par conséquent la fameuse thèse de "l'East Side Story", qui supposait que le clade des Hominidés, et sa posture bipède caractéristique, s'étaient développés en Afrique de l'Est, dans la vallée du rift, en relation avec le climat plus sec, et le milieu plus ouvert que dans l'Ouest, qui s'y seraient installés après l'ouverture de ce fossé d'effondrement.
Il convient cependant de signaler qu'une discussion s'est engagée sur la nature exacte de ce fossile : Si ses découvreurs en font l'un des premiers membres de la lignée humaine, d'autres chercheurs (par ailleurs eux-mêmes découvreurs d'un autre fossile ancien, Orrorin tugenensis, voir ci-après) l'interprètent comme un représentant primitif de la lignée des Gorilles. Le débat est engagé, en termes assez vifs·Les arguments avancés par les deux partis ont été présentés dans deux communications du journal Nature, daté du 10 octobre 2002.
Orrorin tugenensis, -6 Ma :
|
|
Ci-contre, l'ensemble du matériel fossile découvert au Kenya, dans la vallée du Rift, en décembre 2000, et attribué à un nouvel Hominidé, âgé d'environ 6 Ma, Orrorin tugenensis. |
NEOGENE
3-PLIOCENE (5,3-1,75 Ma)
L'environnement :
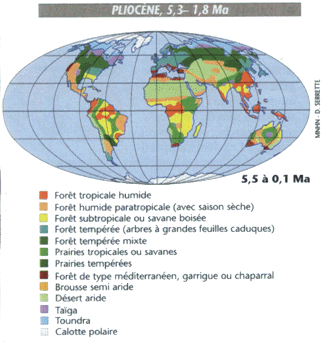
La disposition des continents est désormais très semblable à sa configuration actuelle, la répartition générale des grandes zones climatiques également. Par contre, le Pliocene connaît des modifications fréquentes et souvent abruptes du climat global, qui se poursuivent à l'Holocène, et que l'on reconstitue aujourd'hui grâce aux indicateurs paléoclimatiques à la disposition des chercheurs.

Ajouter un commentaire